
Auf dieser Seite sammle ich zum Lernen und Wiederholen, was ich bei meiner Unterrichts-Vorbereitung an thematischen Vorgaben in Lehrplänen und Schulbüchern sowie inhaltlich in richtigen Fachbüchern, naturwissenschaftlichen Dokumentationen und naturwissenschaftlicher Primärliteratur finde. Was davon glaubwürdig und korrekt ist, versuche ich aufgrund meiner Fachkompetenz als Diplombiologe mit den Hauptfächern Genetik, Biochemie und Organische Chemie zu beurteilen. Zur Veranschaulichung nutze ich mit Creative-Commons-Lizenzen oder als Pulic Domain freigegebene grafische Darstellungen oder stelle selbst her, was ich im World Wide Web nicht finde.
Zu diesem Selbstlern-Hypertext habe ich ein kleines "Arbeitsblatt" mit Aufgaben erstellt.
|
Was sieht der Kernlehrplan für die Sekundarstufe 1 zum Thema Genetik vor? |
|---|---|
|
Grundlagen der klassischen Genetik |
|
die mendelschen Regeln 1 und 2 |
|
die 3. Mendelsche Regel |
|
Mendelsche Regeln am Beispiel der Wellensittich-Farben |
|
Zellkerne als Bibliotheken eukaryotischer Zellen |
|
Chromosomen sind die Kochbücher des Lebens. |
|
die Chromosomen des Menschen |
|
Chromosomen werden kopiert und vererbt |
|
Verdoppelt wird in Chromosomen hauptsächlich die DNA. |
|
ein Erbgang ohne Crossing-over |
|
die Strukturen von Nukleotiden und DNA |
|
von Aminosäuren zu Proteinen |
|
Wofür brauchen Lebewesen Baupläne? |
|
Lebewesen sind ständig neu materialisierte Information. |
|
Biopolymere aus unterschiedlichen Monomeren können Informationen in Sequenzen enthalten. |
|
DNA in Chromosomen als Träger der Erbinformation |
|
Vom Gen zum Genprodukt und zur Eigenschaft |
|
Lernkasten Genetik und Evolution |
|
der Zellzyklus |
|
Mitose |
|
Meiose |
|
Fachbegriffe zum Thema Vererbung |
|
weiterführende Quellen |
|
Was sieht der Kernlehrplan für die Sekundarstufe 1 zum Thema Genetik vor?
|
|---|
Der Biologie-Kernlehrplan verlangt in Kapitel 4 (Inhaltsfelder und fachliche Kontexte für das Fach Biologie) für alle G8-Gymnasien in NRW verbindlich, dass im Verlauf der Jahrgänge 7-9 unter anderem das Inhaltsfeld Grundlagen der Vererbung (Dominante/rezessive/kodominante Vererbung, Erbanlagen, Chromosomen, Genotypische Geschlechtsbestimmung und Veränderung des Erbguts) vermittelt werden muss.
Als nicht verbindliche Fachliche Kontexte schlägt der Biologie-Kernlehrplan dazu folgende Themen vor: Gene - Bauanleitung für Lebewesen (Gene - Puzzle des Lebens, genetische Familienberatung).
Die Biologie-Fachkonferenzen einzelner Schulen können mit dem schulinternen Curriculum beschließen, das Thema Genetik schon im zweiten Halbjahr der Jahrgangsstufe 7 vor dem Thema Evolution zu erarbeiten. Am Ende der 7. Klasse sollen dann die Lernenden Chromosomen als Träger der genetischen Information und deren Rolle bei der Zellteilung sowie vereinfacht den Vorgang der Umsetzung vom Gen zum Merkmal an einem Beispiel (Blütenfarbe, Haarfarbe) beschreiben können.
Grundlagen der klassischen Genetik
|
|---|
Ursächlich für unterschiedliches Aussehen verschiedener Spezies sind etwas unterschiedliche Varianten ihrer Proteine. Und die Ursache für die Unterschiede zwischen verschiedenen Varianten eines Proteins sind Unterschiede zwischen ihren Bauplänen, also den Genen. Genauer gesagt sind es Varianten (Allele) des selben Gens.
Menschen erben also ihre Gene von ihren Eltern. Normalerweise haben Frauen von allen und Männer von fast allen ihren Genen zwei Kopien - eine vom Vater und eine von der Mutter. Unsere Eigenschaften sind deshalb eine Mischung der Eigenschaften der jeweiligen Mütter und Väter. Dabei können die Einflüsse der mütterlichen und väterlichen Genvarianten (Allele) gleich oder unterschiedlich groß sein. Kommt von einem Elternteil eine intakte Genvariante und vom anderen Elternteil ein funktionsloses Allel, dann setzt sich das intakte durch. Man nennt es dominant. Defekte Allele, die keine funktionsfähigen Proteine codieren, heißen rezessiv.
die mendelschen Regeln 1 und 2
|
|---|
| Schema eines dominant-rezessiven Erbgangs |
|---|
 |
| Magnus Manske, public domain |
| Das Schema veranschaulicht die ersten beiden mendelschen Regeln anhand eines dominant-rezessiven Erbgangs, wobei das Allel für eine rote Blütenfarbe dominant ist. Ein defektes Allel dieses Gens führt zu einer weißen Blütenfarbe. Dieses Merkmal ist jedoch rezessiv, weil das defekte Allel durch das intakte ersetzt werden kann. (1) Elterngeneration mit reinerbigen Anlagen (w/w oder R/R). (2) F1-Generation: Alle Individuen sehen gleich aus, die dominante rote Erbanlage setzt sich gegen die rezessive weiße durch. (3) F2-Generation: Dominante (rot) und rezessive (weiße) Erscheinungsformen zeigen ein Verhältnis von 3:1. |
Sind beide Allele gleich dominant, dann führt das zu folgendem Erbgang.
| Schema eines intermediären Erbgangs |
|---|
 |
| Magnus Manske, public domain |
| Das Schema veranschaulicht die ersten beiden mendelschen Regeln anhand eines intermediären Erbgangs, wobei die Kombination beider Allele eine Mischfarbe egibt. (1) Elterngeneration mit reinerbigen Anlagen (w/w oder R/R). (2) F1-Generation: Die Blütenfarbe aller Individuen ist rosa. (3) F2-Generation: Rote, rosa und weiße Blüten zeigen ein Verhältnis von 1:2:1. |
die 3. Mendelsche Regel
|
|---|
Das folgende Schema veranschaulicht die dritte mendelsche Regel mit zwei unabhängig voneinander vererbten Merkmalen. Unten in der Tabelle sieht man, wie sich die Merkmale auswirken. Man nennt diese äußerlich erkennbaren Merkmale den Phänotyp. Phänotypisch unterscheidbar sind die Merkmale Farbe (grün oder gelb) und Form (glatt oder wellig) der Bohnen. Die Tabelle zeigt für den Fall zweier hinsichtlich beider Merkmale mischerbiger Eltern die relativen Häufigkeiten der Merkmalskominationen unter den Nachkommen.
Über der Tabelle und mit den Buchstaben auch in ihr zeigt das Schema die genotypische Erklärung, also die nicht direkt sichtbare Ebene der Gene. Man sieht Zellen mit größeren und etwas kleineren Chromosomen. Die größeren Chromosomen enthalten ein Gen, welches die Farbe der Bohne beeinflusst. Die kleineren Chromosomen enthalten ein Gen, welches die Form der Bohne beeinflusst. Von jedem der beiden Gene gibt es zwei Varianten (Allele), wobei jeweils die groß geschriebene dominant ist und sich gegenüber dem klein geschriebenen rezessiven Allel durchsetzt. Das Allel R bewirkt eine grüne Färbung. Das rezessive Allel r bewirkt eine gelbe Färbung, sofern es nicht durch ein Allel R dominiert wird. Analog lässt das dominante Allel Y die Bohne wellig wachsen, während das rezessive Allel y für eine glatte Bohne sorgt, wenn es nicht von Y dominiert wird.
Gemäß der dritten mendelschen Regel werden die beiden Merkmale unabhängig voneinander vererbt, weil sie auf verschiedenen Chromosomen untergebracht sind. Lägen die beiden Gene direkt nebeneinander im selben Chromosom, dann würden sie fast nie unabhängig voneinander verberbt.
| Schema zur Unabhängigkeitsregel |
|---|
|
Ganz oben unter der Überschrift Meiose zeigt das Schema zwei (diploide) Körperzellen der Elterngeneration (F1-Generation). Man sieht in jeder Zelle zwei größere und zwei etwas kleinere Chromosomen, von denen jeweils eines von der Mutter und eines vom Vater stammt. Die größeren Chromosomen enthalten ein Gen (R oder r), welches die Farbe der Bohne beeinflusst. Die kleineren Chromosomen enthalten ein Gen (Y oder y), welches die Form der Bohne beeinflusst. Beide Körperzellen sind hinsichtlich beider Gene mischerbig. Das bedeutet, dass jeweils das vom "Großvater" stammende Chromosom ein anderes Allel das von der "Großmutter" stammende enthält. Weil sich jeweils das dominante Allel durchsetzt, sind die Bohnen der in diesem Experiment als Vater und Mutter eingesetzten Elternpflanzen alle grün und wellig. In der zweiten Zeile sieht man, das alle Chromosomen (durch DNA-Replikation) verdoppelt wurden. Die beiden Kopien (Chromatiden) hängen aber (an den Centromeren) noch zusammen. Es folgt die erste meiotische Zellteilung, in welcher jede (der haploiden) Tochterzelle von jedem Chromosomenpaar nur entweder das großväterliche oder das großmütterliche Chromosom erhält. Das Ergebnis dieser ersten meiotischen Zellteilung sieht man in der dritten Zeile mit vier (haploiden) Tochterzellen. Abgeschlossen wird die Bildung von in diesem Fall weiblichen Geschlechtszellen (Eizellen) durch die zweite meiotische Zellteilung. Dabei werden die beiden Kopien (Schwesterchromatiden) jedes Chromosoms voneinander getrennt und jede der Tochterzellen in der vierten Zeile erhält von jedem Chromosom nur eine Kopie (Chromatid). Insgesamt verfügt dadurch jede Geschlechtszelle (Gamete) von jedem Chromosom und im Prinzip auch von jedem Gen nur noch jeweils eine Kopie. Und weil die Körperzellen hinsichtlich beider Gene mischerbig waren, enthalten nun die Geschlechtszellen unterschiedliche Allelkombinationen. |
 |
| Mariana Ruiz Villarreal, gemeinfrei |
|
Unten zeigt die Tabelle alle möglichen Kombinationen der vier Allele, die durch Befruchtung der Eizellen durch genauso entstandene männliche Geschlechtszellen in der Generation der Nachkommen (F2-Generation) entstehen können. |
Werden bei einem Heterozygoten die Genprodukte beider Allele unabhängig voneinander ausgeprägt, spricht man von Kodominanz bzw. von kodominanter Vererbung. Das bekannteste Beispiel für Kodominanz findet sich beim Blutgruppensystem AB0: Die Allele A und B kommen beim Genotyp AB beide zur Ausprägung, so dass auch der Phänotyp AB lautet. Das Gen zur Ausprägung der Blutgruppen des AB0-Systems liegt auf dem Chromosom 9. Beispiele für intermediäre Erbgänge sind die rosafarbenen Blüten von Primeln, Löwenmäulchen oder ursprünglich mittelamerikanischen Wunderblume, weil das Merkmal Weiß auf zwei von Vater und Mutter geerbten defekten Allelen ohne Genprodukt beruht und ein intaktes Allel allein nicht ausreicht, um genügend roten Farbstoff für rote Blüten zu produzieren. Kreuzt man zwei jeweils homozygot weiße und rote Exemplare dieser Blütenpflanzen miteinander, dann findet man in der nächsten Generation nur rosafarbene Blüten.
Mendelsche Regeln am Beispiel der Wellensittich-Farben
|
|---|
Es gibt bei den seit mehr als 170 Jahren als Haustiere gezüchteten Wellensittichen zahlreiche Farbvarianten. Um es nicht unnötig kompliziert zu machen, möchte ich mich darauf beschränken, die Vererbung von zwei an der Farbgebung beteiligten Genen mit jeweils 2 Allelen zu erklären.
Das eine Gen ist ein Rezept für die Produktion eines Proteins, welches normalerweise zur Herstellung des dunklen Farbstoffs Eumelanin führt. Zusammen mit der normalen Struktur der Federn bewirkt dieser Farbstoff eine Blaufärbung. Es gibt aber ein mutiertes Allel (b) dieses Gens, das zu einem defekten Protein führt. Dieses nicht funktionsfähige Protein kann keinen Beitrag zur Herstellung des Farbstoffs leisten. Wenn der Vogel auch von seinem anderen Elternteil kein intaktes Allel (B) dieses Gens geerbt hat, dann kann er gar keinen Farbstoff für die Blaufärbung produzieren.
Das andere Gen codiert ein Protein, welches für die Produktion des gelben Farbstoffes Psittacin benötigt wird. Auch von diesem Gen gibt es außer dem funktionsfähigen (G) noch ein funktionsunfähiges mutiertes Allel (g).
Betrachten wir mit diesem Vorwissen die Kreuzung eines reinerbig grünen (BBGG) mit einem reinerbig weißen (bbgg) Wellensittichs, dann enthalten alle Geschlechtszellen des einen Elternteils die beiden dominanten Allele B und G, während alle Gameten des anderen Tieres die rezessiven Allele b und g enthalten. Bei den Nachkommen ergeben sich folgende Kombinationsmöglichkeiten:
| F1 | bg | bg |
| BG | BbGg | BbGg |
| BG | BbGg | BbGg |
Alle Nachkommen in der F1-Generation sind genotypisch und phänotypisch gleich. Alle sind grün. Sie sind aber auch alle mischerbig. Deshalb produzieren sie im Hinblick auf diese beiden Gene 4 unterschiedliche Sorten von Geschlechtszellen: BG, Bg, bG und bg.
| F2 | BG | Bg | bG | bg |
| BG | BBGG | BBGg | BbGG | BbGg |
| Bg | BBGg | BBgg | BbGg | Bbgg |
| bG | BbGG | BbGg | bbGG | bbGg |
| bg | BbGg | Bbgg | bbGg | bbgg |
In der F2-Generation findet man also die Farben Grün, Blau, Gelb und Weiß im Verhältnis: 9:3:3:1.
Zellkerne als Bibliotheken eukaryotischer Zellen
|
|---|
Das Kapitel: "Der Aufbau einer tierischen Zelle" im "Lerntext Organellen" zeigt den Zellkern und andere wichtige Organellen einer tierischen Zelle.
Informationen zu den Strukturen und Funktionen von Zellkern (Nukleus) und Nukleoli sowie zu den Kernporen findet man im Kapitel Zellkerne als Bibliotheken eukaryotischer Zellen des Lerntextes Organellen.
| Löse ausschließlich mit Hilfe des Lerntext-Kapitels Zellkerne als Bibliotheken eukaryotischer Zellen die Aufgaben des Lernmoduls Zellkerne als Bibliotheken eukaryotischer Zellen! |
Nützlich für das Verständnis der Rolle des Zellkerns ist eine genauere Betrachtung der Metamorphose.
| Löse ausschließlich mit Hilfe meiner kritischen Zusammenfassung der Fernsehdokumentation Metamorphose die Aufgaben des Lernmoduls Metamorphose! |
Chromosomen sind die Kochbücher des Lebens.
|
|---|
| Löse ausschließlich mit Hilfe der folgenden 5 Kapitel die Aufgaben des Lernmoduls Chromosomen! |
Jeder der beiden Baupläne im Zellkern einer menschlichen Zelle ist etwa so lang wie ein Mensch und wäre damit so unhandlich wie ein zwei Meter dickes Lexikon. Als lange vor Wikipedia große Lexika wie der Brockhaus oder die Encyclopædia Britannica mit Zigtausend Seiten zu dick und unhandlich wurden, da teilte man sie auf in mehrere kleinere Bücher, die sogenannten Bände. Ganz ähnlich ist das Chromatin der großen und komplexen eukaryotischen Zellen unterteilt in Chromosomen.
Ohne die gesamte Rezeptesammlung (Genom) mit Tausenden Rezepten (Genen) kann keine Zelle lange leben. Die Chromosomen in einem Zellkern sind unterschiedlich groß und können Dutzende bis Tausende Gene (Bauanleitungen) für die Herstellung von Eiweißen und RNAs enthalten.
| Schema eines Bakteriums | |
|---|---|
 |
|
| Mariana Ruiz Villarreal, public domain | |
Auch Viren und Prokaryoten (Bakterien und Archäen) besitzen Chromosomen, aber sie verpacken ihr Erbmaterial nicht in Zellkernen. Bakterien können mit einem einzigen Chromosom auskommen, weil sie nur wenige Tausend Eiweiß-Rezepte (Gene) besitzen. Außerdem sind Bakterien-Zellen so klein, dass ihre Chromosomen von jedem Punkt der Zelle aus leicht erreichbar sind. Vielleicht müssen sie deshalb nicht in einem Zellkern konzentriert werden. Angesichts der häufigen Zellteilungen wäre aber auch das ständige Auf- und Abbauen eines Zellkerns für Bakterien extrem aufwändig.
Der Lerntext Endosymbionten-Theorie erklärt, wie sich Biologen heute die Entstehung der ersten eukaryotischen Zelle aus den beiden Arten von Prokaryoten (Bakterien und Archäen) vorstellen.
die Chromosomen des Menschen
|
|---|
Insgesamt fand man in den menschlichen Chromosomen Gene für rund 23.000 Proteine sowie sehr viele Kopiervorlagen für verschiedene Arten von RNAs, die als Bausteine und Werkzeuge unserer Zellen dienen.
Menschliche Zellkerne enthalten in normalen Körperzellen 46 Chromosomen (23 vom Vater und 23 von der Mutter). Zellkerne menschlicher Geschlechtszellen (Samen- und Eizellen) enthalten nur 23 Chromosomen. Die Geschlechtszellen eines Menschen enthalten unterschiedliche Mischungen der Baupläne seiner beiden Eltern.
Ein Mensch entsteht, wenn sich eine väterliche (Spermium) und eine mütterliche (Eizelle) Geschlechtszelle zu einer befruchteten Eizelle (Zygote) vereinigen. Der Zellkern der Eizelle enthielt 23 Chromosomen, bei denen man zwischen 22 Autosomen und einem Gonosom unterscheidet. Das Gonosom der Eizelle ist immer ein relativ großes X-Chromosom. Auch der Zellkern des Spermiums enthielt 23 Chromosomen, aber sein Gonosom kann entweder ein relativ großes X-Chromosom oder ein nicht einmal halb so großes Y-Chromosom sein. Diesen Unterschied zwischen den menschlichen Gonosomen (Geschlechtschromosomen) sieht man im folgenden elektronenmikroskopischen Bild. Die 23 Chromosomen der Geschlechtszellen nennt man den einfachen oder haploiden Chromosomensatz und auch alle Zellen mit dem einfachen Chromosomensatz heißen haploid.
| die Geschlechtschromosomen X&Y des Menschen |
|---|
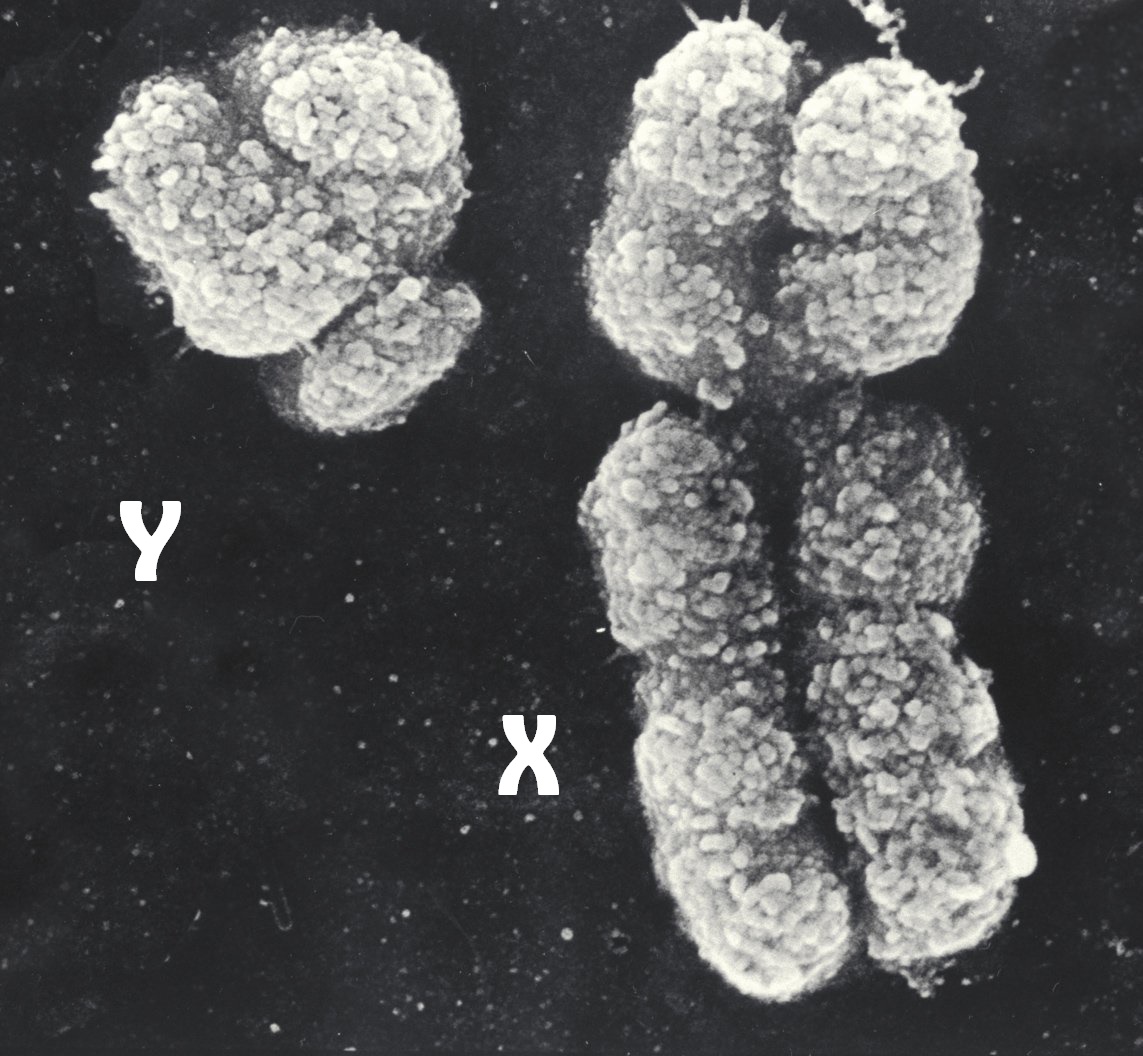 |
| Geo-Science-International, CC BY-SA 4.0 |
| Man sieht im elektronenmikroskopischen Bild zwei menschliche Geschlechtschromosomen in ihrer Transport-optimierten, maximal kondensierten Form. Jedes dieser Chromosomen ist ein Doppelchromosom oder 2-Chromatiden-Chromosom, weil es aus zwei Chromatide genannten, nahezu identischen Kopien besteht. Die Centromere erkennt man daran, dass an diesen Stellen die beiden Chromatiden eingeschnürt sind. |
Nach der Vereinigung der Geschlechtszellen enthält die Zygote 23 mütterliche und 23 väterliche, also insgesamt 46 Chromosomen. Man spricht nun von einem doppelten oder diploiden Chromosomensatz und nennt auch den neuen Zellkern sowie die ganze Zygote diploid. Enthält der Zellkern der Zygote zwei X-Chromosomen, dann ist sie rein genetisch betrachtet weiblich. Enthält aber der Zellkern der Zygote ein X- und ein Y-Chromosom, dann kann sich der Embryo männlich entwickeln, falls alles gut geht. Weil also die Gonosomen das Geschlecht eines Menschen bestimmen, nennt man sie auch Geschlechtschromosomen. Während sich die Gonosomen unterscheiden können, sind die 22 mütterlichen Autosomen genauso groß wie die 22 väterlichen. Die 44 Autosomen bilden 22 Chromosomenpaare und die beiden sich entsprechenden Chromosomen eines Chromosomenpaares heißen homologe Chromosomen.
| Karyogramm eines Mannes |
|---|
 |
| National Human Genome Research Institute, public domain |
Weil die Zellkerne von Frauen zwei X-Chromosomen enthalten, besitzen sie mehr DNA (und Allele) als Männer. Weil aber das Y-Chromosom dem X-Chromosom fehlende, einzigartige Gene enthält, besitzen Männer mehr Gene als Frauen. Wenn in menschlichen Geschlechtsorganen (Eierstöcken oder Hoden) Geschlechtszellen entstehen, dann reduziert eine Meiose genannte spezielle Art der Zellteilung die Zahl der Chromosomen auf nur noch 23. Dadurch wird verhindert, dass die Zellen der Kinder doppelt soviele Chromosomen wie die ihrer Eltern enthalten.
Chromosomen werden kopiert und vererbt
|
|---|
Im Gegensatz zu manchen kochenden Menschen schreiben Zellen ihre Rezeptbücher (Genome) nicht einfach selbst. Genome werden von Mutterzellen an ihre beiden Tochterzellen vererbt oder die beiden Genome einer männlichen und einer weiblichen Geschlechtszelle vereinigen sich zum neuen Genom einer befruchteten Eizelle.
Bei jeder Zellteilung muss jede der beiden Tochterzellen einen eigenen Zellkern mit einer Kopie des kompletten Bauplans (Genoms) erhalten. Deshalb geht jeder Zellteilung eine Kernteilung (Mitose) voraus.
Die Kernteilung darf aber nicht dazu führen, dass sich die beiden neuen Zellkerne die Chromosomen teilen müssen. Denn dann hätte jede Tochterzelle nur noch halb soviele Chromosomen wie die Mutterzelle. Jede Tochterzellen muss von jedem Chromosom eine Kopie erhalten. Darum müssen vor jeder Kernteilung alle Chromosomen des Zellkerns der Mutterzelle verdoppelt werden. Diese Verdopplung der Chromosomen nennt man Replikation oder genauer DNA-Replikation.
Die DNA-Replikation erfogt zwischen einer Zellteilung und der nächsten Kernteilung in der sogenannten S-Phase (Synthese-Phase).
Die beiden Kopien eines Chromosoms heißen Chromatiden. Durch die DNA-Replikation entsteht aus einem 1-Chromatid-Chromosom ein 2-Chromatiden-Chromosom (Doppelchromosom). Aber die beiden Chromatiden bleiben zunächst an einer Centromer genannten Stelle miteinander verbunden. Nach der DNA-Replikation kann die Kernteilung erfolgen.
Solange zwischen den Kernteilungen der Zellkern existiert, bleiben die Chromosomen eukaryotischer Zellen in ihm eingeschlossen. Die folgende Animation zeigt, wie zunächst die 1-Chromatid-Chromosomen durch die DNA-Replikation zu 2-Chromatiden-Chromosomen verdoppelt werden. Danach kommt es zur Kernteilung (Mitose). Schließlich teilt sich die eukaryotische Mutterzelle in zwei Tochterzellen.
Verdoppelt wird in Chromosomen hauptsächlich die DNA
|
|---|
Der wichtigste Bestandteil eines Chromosoms ist ein extrem langes Molekül namens Desoxyribonukleinsäure (DNS), das wie eine spiralig verdrehte Strickleiter aussieht. Auf Englisch nennt man es Desoxyribonucleic Acid. Daher kommt die international übliche Abkürzung DNA. In den Chromosomen der Eukaryoten wird die DNA von Eiweißen kontrolliert und immer in die gerade passende Form gebracht.
Das folgende Schema deutet an, wie in Chromosomen die DNA um sogenannte Histon-Proteine gewickelt ist und wie massiv die normalerweise extrem langen und dünnen Chromosomen zu Beginn (in der Prophase) der Mitose zu dicken, kurzen und dadurch transportfähigen Knubbeln aufgewickelt und gerafft (kondensiert) werden.
| Schema zur Kondensierung eines Chromosoms |
|---|
 |
| NIH, Armin Kübelbeck, CC BY-SA 3.0 |
Die Verdopplung eines Chromosoms erfolgt hauptsächlich durch das DNA-Replikation genannte Kopieren oder Verdoppeln der darin enthaltenen DNA. Im Detail wird die DNA-Replikation im Lerntext Zellbiologie erklärt. Während der DNA-Replikation müssen die Chromosomen ganz lang und entknäuelt sein, weil sie sonst nicht kopiert werden könnten. Aber während der Kernteilung müssen sie transportiert werden. Und es wäre extrem aufwändig, in einer winzigen Zelle ein vielfach längeres Chromosom vollständig in eine Ecke zu ziehen. Zu Beginn der Mitose muss daher jedes Chromosom ganz eng zusammen geknäuelt werden. In dieser kondensierten Transportform sieht man die Chromosomen oben im oberen Drittel des Schemas.
Insgesamt enthalten die 46 Chromosomen in den Zellkernen menschlicher Körperzellen gut sechseinhalb Milliarden Basenpaare, die vor einer Mitose alle kopiert werden müssen. Trotz dieser enormen Menge wäre es vermutlich möglich, perfekte Kopien zu machen. Das tun Zellen aber nicht. Ihre Kopierwerkzeuge arbeiten nicht vollkommen perfekt, weil alle Lebewesen mit perfekt arbeitenden Kopierwerkzeugen ausgestorben sind. Man nennt die zufällig auftretenden Kopierfehler Mutationen und ohne Mutationen könnten sich Spezies nicht an veränderte Umwelten anpassen.
ein Erbgang ohne Crossing-over
|
|---|

die Strukturen von Nukleotiden und DNA
|
|---|
Der Lerntext Biomoleküle liefert Informationen zu den Themen:
von Aminosäuren zu Proteinen
|
|---|
Der Lerntext Biomoleküle liefert auch Informationen zu den Themen:
Wofür brauchen Lebewesen Baupläne?
|
|---|
In der menschlichen Arbeitswelt werden Baupläne verwendet, wenn wir komplizierte Dinge bauen. Und in der Küche benutzen wir Rezepte, wenn wir komplizierte Kuchen backen.
Lebewesen sind extrem komplexe, offene Systeme, in denen sehr viele Sorten von Biomolekülen zeitlich, räumlich und funktionell exakt koordiniert zusammen arbeiten. Tiere und Pilze nehmen einige (essenzielle) dieser Biomoleküle mit der Nahrung auf. Alle Lebewesen stellen aber zumindest die meisten ihrer Biomoleküle mit Hilfe von Enzymen selbst her. Enzyme sind Biokatalysatoren, also von Lebewesen produzierte Katalysatoren. Für diese Produktion ihrer eigenen Enzyme und aller anderen Proteine und RNAs benötigen Lebewesen Baupläne, Bauanleitungen oder Rezepte.
Umgangssprachlich sprechen wir von einem Bauplan, wenn wir die Bauanleitung für ein einzelndes Protein oder eine RNA meinen. Bauplan nennen wir aber auch die Summe aller Bauanleitungen, die gemeinam den Bauplan für den gesamten Organismus darstellen. Um zwischen diesen beiden Ebenen leichter unterscheiden zu können, haben wir die Fachbegriffe Genen für die einzelne Bauanleitung und Genom für die Summe aller Bauanleitungen.
Lebewesen sind ständig neu materialisierte Information.
|
|---|
Was ein Lebewesen als Individuum ausmacht, das sind sein Bauplan und die Gesamtheit aller Informationen, die auf vielen Ebenen in seinen Strukturen stecken. Denn biologische Informationen können viel dauerhafter als ihre Informationsträger (Strukturen) sein und Strukturen viel unveränderlicher als Zusammensetzung ihrer Bestandteile.
Was ein Lebewesen wirklich ausmacht, sind daher nicht seine Atome und Moleküle, sondern die in ihm steckenden Informationen. So stecken beispielsweise unsere Gedächtnisinhalte in Strukturen und Verknüpfungen von Nervenzellen. Und obwohl bei jeder Zellteilung die Hälfte aller Atome in der DNA im Zellkern einer Zelle ersetzt wird, bleiben die in den DNA-Sequenzen (Nukleotid-Reihenfolgen) steckenden Informationen lebenslang nahezu unverändert erhalten. Diese von unseren Eltern ererbten Baupläne (Genome) sind gigantische Rezeptbücher mit Zigtausenden Rezepten (Genen) für Proteine (Eiweiße) und RNAs.
Biopolymere aus unterschiedlichen Monomeren können Informationen in Sequenzen enthalten.
|
|---|
Wir verwenden traditionell Papier, um auf diesem organischen Material als Informationsträger mit Stiften Informationen aufzuschreiben oder zu zeichnen. Der ebenfalls aus organischem Material bestehende Informationsträger in unseren Zellen ist die DNA. DNA und Proteine sowie die großen RNAs und Kohlenhydrate sind Makromoleküle, weil jedes ihrer Moleküle aus weit mehr als 1000 Atomen besteht. Und sie sind Polymere, weil sie aus vielen kleinen Grundbausteinen, den sogenannten Monomeren aufgebaut sind. Poly bedeutet viele und Mono bedeutet 1. Biopolymere heißen sie, weil sie von Lebewesen produziert werden. Im Gegensatz zu chemisch hergestellten Polymeren bestehen die Biopolymere Nukleinsäuren und Proteine aus unterschiedlichen Monomeren. Die Reihenfolge (Sequenz) der Monomere beeinflusst die Eigenschaften der Biopolymere.
| Monomere, Oligomere und Polymere | |
|---|---|
 |
Monomer nennt man einen Baustein, eine Untereinheit oder eine Perle, die eine lange Kette bildet. Beispiele für Monomere unter den Biomolekülen sind Aminosäuren und Nukleotide. Oligomer nennt man eine kurze Kette aus mehreren Monomeren. Beispiele dafür sind Oligonukleotide und die Peptide, die beispielsweise bei der Vorverdauung von Proteinen im Magen entstehen. Polymer nennt man eine lange Kette aus vielen Monomeren. Beispiele sind Ketten aus vielen Perlen, Proteine aus vielen Aminosäuren oder Nukleinsäuren (DNA oder RNA) aus vielen Nukleotiden. |
| Roland Heynkes, CC BY-SA-4.0 | |
Die Bedeutung (Information) eines Wortes steckt nicht in Informationsträgern wie Papier und Tinte oder in seinen Buchstaben, sondern in der Reihenfolge (Sequenz) der Buchstaben auf dem Papier. Auch genetische Informationen (Erbinformationen) stecken nicht einfach im Material des Biopolymers DNA oder in deren Nukleotide genannten Monomeren. Weil Nukleinsäuren und Proteine im Gegensatz zu Kohlenhydraten unverzweigt sind, steckt bei ihnen die Information einfach in der Sequenz der Monomere. Die Erbinformationen stecken in den Sequenzen der DNA-Nukleotide (genauer: Desoxyribonukleotide). Die DNA ist allerdings eher mit einem Satz oder einem ganzen Buch als mit einem Wort zu vergleichen, weil die Nukleotidsequenz jedes DNA-Moleküls mehrere bis Tausende Rezepte (Gene) für die Herstellung von Eiweißen (Proteinen) oder RNAs enthält.
Menschliche Sprachen haben viele Buchstaben, aber digital speichern wir Texte, Bilder und Filme mit nur 2 unterschiedlichen Zuständen wie 0 und 1, An oder Aus, Loch oder kein Loch. In der DNA gibt es nur 4 verschiedene Sorten von Nukleotiden, deren Namen man mit A, C, G und T abkürzt. Unsere Erbinformationen mit den Bauanleitungen für alle unsere RNAs und Proteine sind also aufgeschrieben (codiert) in der DNA mit einem Alphabet aus nur 4 Buchstaben.
Prinzipiell reichen 4 Buchstaben für die Codierung der Aminosäure-Sequenz eines Proteins durch die Nukleotid-Sequenz einer Nukleinsäure. Allerdings stehen die nur 4 Sorten von Nukleotiden als Monomere der DNA der deutlich größeren Anzahl von 20 unterschiedlichen Aminosäuren als Monomere menschlicher Proteine gegenüber. Um 20 verschiedene Aminosäuren mit nur 4 unterschiedlichen Nukleotiden codieren zu können, müssen jeweils 3 Nukleotide zu einem Codon zusammengefasst werden, denn Codons aus nur 2 Nukleotiden könnten nur 4x4=16 verschiedene Aminosäuren codieren. Das folgende Schema soll leichter nachvollziehbar machen, wie Aminosäuresequenzen durch Nukleotidsequenzen in DNA codiert werden.
| Gene codieren Proteine |
|---|
 |
Ebenfalls in die DNA geschriebene regulatorische Sequenzen binden Botenstoffe aus der eigenen und vielen anderen Zellen, welche das Kopieren einzelner Bauanleitungen (Gene) fördern oder verhindern.
DNA in Chromosomen als Träger der Erbinformation
|
|---|
Doppelsträngige DNA ist ein unglaublich langes Riesenmolekül, das wie eine spiralig verdrehte Strickleiter aussieht. Zusammen mit zahlreichen Proteinen bildet die DNA Chromosomen, die einen (1-Chromatid-Chromosom) oder zwei (2-Chromatiden-Chromosom oder Doppelchromosom) DNA-Doppelstränge enthalten können. Vergleicht man die Gesamtheit der im Zellkern gespeicherten Erbinformationen mit einem großen Lexikon, dann entsprechen die Chromosomen den einzelnen Bänden, in welche das Lexikon aufgeteilt wurde, damit es handhabbar bleibt.
Vom Gen zum Genprodukt und zur Eigenschaft
|
|---|
Der in DNA gespeicherte Bauplan (Genom) befindet sich bei noch teilungsfähigen eukaryotischen Zelle im Zellkern. Und wie die wertvollsten Bücher einer Bibliothek nur fotografiert oder photokopiert und nicht ausgeliehen werden, so verlässt auch die DNA nicht den Zellkern. Braucht eine Zelle ein Rezept (Gene) für ein bestimmtes Protein, dann werden im Zellkern von dem entsprechenden Gen in einem Transkription genannten Prozess Umschriften (Kopien auf den etwas anderen Informationsträger RNA) gemacht. Weil eine Sorte von ihnen wie Boten wirken, nennt man diese speziellen RNAs Boten-RNA oder kurz mRNA. Sie binden im Cytoplasma an Ribosomen, die mit Hilfe sogenannter tRNAs fähig sind, die von den mRNAs codierten Proteine zu synthetisieren. Diese Übersetzung einer Nukleotidsequenz in eine Aminosäuresequenz nennt man Translation. (Wie Transkription und Translation funktionieren, können Interessierte im Lerntext Genetik für die gymnasiale Qualifikationsphase nachlesen.)
Proteine können einfach Bausteine unseres Körpers, Rezeptoren, Antikörper, Kanäle, Pumpen oder Enzyme sein. Alle diese Proteine beeinflussen das Aussehen oder die Fähigkeiten eines Lebewesens und teilweise unterscheiden sie sich von Spezies zu Spezies und von Rasse zu Rasse.
Lernkasten Genetik und Evolution
|
|---|
Die rote Umrandung des folgenden Abschnittes soll andeuten, dass die darin enthaltenen 8 Sätze kompakt zusammenfassen, was Lernende eigentlich schon am Ende der Sekundarstufe 1 über Genetik und Evolution wissen sollten. Spätestens in der Erprobungsstufe müssen diese 8 Sätze vollständig verstanden und sinngemäß gewusst werden.
|
der Zellzyklus
|
|---|
Nach der DNA-Replikation muss nur noch dafür gesorgt werden, dass jede Tochterzelle eine Kopie erhält. Bei sich alle 20 Minuten teilenden Bakterien werden allerdings die gerade als Ergebnis einer noch laufenden DNA-Replikation entstehenden Tochterchromosomen schon wieder einer weiteren Replikation unterzogen, damit die Vermehrung der Bakterien-Chromosomen mit der extremen Zellteilungsrate mithalten kann.
Bei Eukaryoten ist die Sache komplizierter. Im vollständigen Zellzyklus der Eukaryoten unterscheidet man meistens nicht nur zwischen den beiden Phasen Wachstum (Interphase) und Zellteilung. Sinnvoll ist zunächst innerhalb der Zellteilung eine Unterscheidung zwischen Kernteilung (Mitose) und Plasmateilung (Cytokinese), weil auf eine Kernteilung nicht immer eine Plasmateilung folgt. Die normale Mitose wird traditionell in Prophase, (Prometaphase), Metaphase, Anaphase und Telophase unterteilt, während man in der Wachstumsphase zwischen G1-, S- und G2-Phase unterscheidet. In der G1-Phase wächst eine Zelle ungefähr zur ursprünglichen Größe heran. Dann folgt die S-Phase, in der die DNA-Replikation, stattfindet. Ihr folgt die G2-Phase als Pause zwischen der Verdopplung des Bauplans und der Kernteilung. Es folgt die Kernteilung (Mitose) als Voraussetzung für die Plasmateilung (Cytokinese). Um den Zellzyklus abzuschließen, muss nun nur noch die (Cytokinese) erfolgen, die aus einer großen zwei kleine Zellen macht. Dazu werden in der Äquatorialebene zwei neue Zellmembranen und bei Pflanzen zusätzlich zwei Zellwände aufgebaut.
| Der Zellzyklus bei Eukaryoten | |
 |
|
Mit dem für meinen Geschmack entschieden zu teuren Programm: "Adobe After Effect" hat Jessica Scheiderer im Rahmen ihrer Bachelor-Arbeit ein tolles Video zur Erklärung der Mitose im Zellzyklus produziert.
Transkription und DNA-Replikation funktionieren am besten, wenn die DNA möglichst entspannt ist. Schließlich müssen die DNA-Doppelstränge getrennt und um neue Gegenstränge ergänzt werden. Man nennt die Phase der DNA-Replikation auch Synthese-Phase und in ihr kann man die Chromosomen lichtmikroskopisch nicht sehen. Nach der DNA-Replikation enthält jedes Chromosom zwei DNA-Doppelstränge. Jeder der beiden DNA-Doppelstränge ist um spezielle Proteine gewickelt und man nennt diese aus Proteinen und DNA bestehende Struktur ein Chromatid. An einer Centromer genannten Stelle sind die beiden Chromatiden eines 2-Chromatiden-Chromosoms miteinander verbunden. Damit jeder der beiden neuen Zellkerne von jedem Chromosom eine Kopie erhält, müssen in jedem Chromosom die beiden Chromatiden voneinander getrennt und in unterschiedliche Hälften der Zelle gezogen werden. Man kann aber sehr lange DNA-Fäden sehr schlecht transportieren. Vor dem Transport werden deshalb die Chromatiden extrem dicht verknäuelt, sodass sie dick, kurz und dadurch auch sichtbar werden. Danach werden zunächst alle Chromosomen in eine Ebene in der Zellmitte transportiert. Dann werden die beiden Chromatiden jedes Chromosoms getrennt und in entgegengesetzte Richtungen gezogen. Schließlich bildet das endoplasmatische Retikulum in jeder Zellhälfte eine neue Kernhülle um die jetzt nur noch aus jeweils einem Chromatid bestehenden Chromosomen. Diese entknäueln sich jetzt wieder und werden dadurch wieder quasi unsichtbar.
Mitose
|
|---|
Mitose oder Kernteilung nennen Biologen den Teil des Zellzyklusses, der von der Prophase bis zur vollendeten Telophase reicht.
Die bereits während der DNA-Replikation verdoppelten Centrosomen wandern während der Prophase weiter zu den beiden Zellpolen. Zwischen den beiden Centrosomen entwickelt sich der Spindelapparat.
In der Prophase kondensieren (ballen sich zusammen) die Chromosomen zu dicken, aber kurzen und damit transportfähigen Einheiten. In dieser Phase sieht man, dass jedes Chromosom aus zwei Schwesterchromatiden besteht. Das sind zwei nahezu identische Kopien des selben Chromosoms, die noch am Centromer aneinander gebunden sind und deshalb beide als Bestandteile eine 2-Chromatiden-Chromosoms bezeichnet werden.
| Schema der Prophase einer tierischen Zelle |

|
| anonymer Amerikaner, Roland Heynkes, CC BY-SA-4.0 |
In der Prometaphase wird die Kernhülle aufgelöst und die Mikrotubuli des Spindelapparates wachsen bis zu den Centromeren der Chromosomen.
| Schema der Prometaphase einer tierischen Zelle |

|
| anonymer Amerikaner, Roland Heynkes, CC BY-SA-4.0 |
In der Metaphase werden die Chromosomen vom Spindelapparat alle in eine Ebene in der Mitte der Zelle bugsiert, die man Äquatorialebene nennt.
| Schema der Metaphase einer tierischen Zelle |

|
| anonymer Amerikaner, Roland Heynkes, CC BY-SA-4.0 |
In der Anaphase zieht der Spindelapparat die beiden Schwesterchromatiden der Chromosomenen auseinander bis an die gegenüber liegenden Pole der Zelle. Weit von einander entfernt liegt nun in beiden Hälften einer Zelle von jedem Chromosom jeweils ein Chromatid und man bezeichnet deshalb die Chromosomen von nun an bis zur nächsten S-Phase als 1-Chromatid-Chromosomen.
| Schema der Anaphase einer tierischen Zelle |

|
| anonymer Amerikaner, Roland Heynkes, CC BY-SA-4.0 |
In der Telophase löst sich der Spindelapparat auf, an den beiden Zellpolen bilden sich um die 1-Chromatid-Chromosomen herum neue Kernhüllen und in diesen dekondensieren die Chromosomen wieder. Das bedeutet, dass sie wieder ganz lang und dünn werden, sodass man keine einzelnen Chromosomen mehr erkennen kann. Man sieht nur noch eine einzige Masse, die aufgrund ihrer Färbkarkeit Chromatin nennt.
| Schema der Telophase einer tierischen Zelle |

|
| anonymer Amerikaner, Roland Heynkes, CC BY-SA-4.0 |
Meiose
|
|---|
Meiose oder Reduktionsteilung nennt man eine besondere Form der Zellteilung, bei der in zwei Teilungsschritten aus einer diploiden Zelle vier haploide Geschlechtszellen entstehen. Im ersten Schritt werden die Chromosomenpaare getrennt und im zweiten die Schwesterchromatiden. Diese Reduktionsteilung ist bei der Entstehung von Gameten notwendig, weil sich sonst bei jeder Befruchtung die Zahl der Chromosomen verdoppeln würde. Die Verteilung der ursprünglich mütterlichen und väterlichen Chromosomen erfolgt zufällig und zusätzlich kommt es noch zum Austausch von Chromosomenteilen, sodaß jede Geschlechtszelle eine einzigartige Mischung der elterlichen Gene enthält. Die meisten Quellen gehen überhaupt nicht auf diese Frage ein, aber der BioCoach des Pearson Verlages erklärt, dass die 2-Chromatiden-Chromosomen in vielen, aber nicht in allen Zellen nach der ersten meiotischen Kernteilung dekondensieren.
Die Animation funktioniert unter MS-Windows mit den Browsern Firefox, Opera und Google Chrome, während der Internet Explorer von Microsoft die SMIL-Anweisungen immer noch nicht versteht. Auf dem iPad muss die Seite vor jeder Wiederholung neu aufgebaut werden.
Schema der Stadien der Meiose mit Crossing over 
|
|---|
|
Die Meiose kommt nur bei Lebewesen vor, deren noch lebende Zellen einen Zellkern haben. Diese Lebewesen heißen Eukaryoten. Eukaryoten setzten die Meiose nur für die Herstellung von Geschlechtszellen ein. Der Zweck der Meiose ist die Halbierung des in normalen Körperzellen doppelten (diploiden) Chromosomensatzes. Denn Geschlechtszellen dürfen von jedem Chromosom nur ein Exemplar enthalten, damit sich die Zahl der Chromosomen in den Körperzellen nicht bei jeder sexuellen Fortpflanzung verdoppelt. Die Meiose beginnt mit Chromosomenpaaren aus jeweils zwei Chromosomen, die normalerweise Varianten der gleichen Gene in der selben Reihenfolge enthalten. Sich so entsprechende Chromosomen nennt man homologe Chromosomen. Dabei stammt jeweils ein Chromosom von der Mutter und das andere vom Vater. Und weil schon vor der Meiose jedes Chromosom kopiert oder genauer verdoppelt wurde, besteht jedes Chromosom aus zwei Kopien, die man Chromatiden nennt. Deshalb nennt man die Chromosomen vor der Meiose 2-Chromatiden-Chromosomen. |

|
| anonym, CC BY-SA 4.0 |
|
Das Schema oben beginnt mit drei Paaren homologer Chromosomen mit je zwei Chromatiden. Dabei stammen die blauen Chromosomen vom einen und die roten vom anderen Elternteil. Gelb gezeichnet sind die für den Transport der Chromosomen benötigten Teile des Zytoskeletts. Das Schema unterteilt die Meiose in zwei Zellteilungen (1. Reduktionsteilung, 2. Äquationsteilung genannte normale Mitosen) mit insgesamt acht Phasen:
|
Fachbegriffe zum Thema Vererbung
|
|---|
Die folenden Begriffe werden von Schulbuchautoren gerne benutzt, in den meisten Fällen aber nicht erklärt. Weil das zu einer Diskriminierung von Lernenden mit Migrationshintergrund oder aus nichtakademischen Elternhäusern führt, erkläre ich sie an dieser Stelle.
weiterführende Quellen
|
|---|
Der High-School-Lehrer Paul Andersen hat zahlreiche Lehrvideos produziert, die man bei Bozemanscience und YouTube sehen kann. Zum Thema Genetik findet man von ihm unter anderem:
Das berühmte Massachusetts Institute of Technology (MIT) stellt seit 2002 Tausende Vorlesungen und Kurse kostenlos als sogenannte MIT OpenCourseWare auf seinen eigenen Internetseiten und bei YouTube zur Verfügung.
The Royal Society stellt zahlreiche Lernvideos über YouTube zur Verfügung. Darunter auch viele zum Thema Genetik.
Die Nobelpreisträgerin Elizabeth Blackburn (Department of Biochemistry an der University of California, San Francisco) erklärt in einer dreiteiligen Vorlesung (Part 1: The Roles of Telomeres and Telomerase, Part 2: Telomeres and telomerase in human cells and in cancer und Part 3: Stress, Telomeres and Telomerase in Humans) die Bedeutung der Telomere und Telomerase (Telomeres and telomerase: Their implications in human health an disease). Man findet diese Reihe und weitere Vorträge von Elizabeth Blackburn auch im Biologie-Portal iBiology.
Besonders Interessierte finden bei YouTube drei Genetik-Vorlesungen des Harvard-Professors und Nobelpreisträgers Jack Szostak: Part 1: The Origin of Cellular Life on Earth, Part 2: Protocell Membranes und Part 3: Non-enzymatic Copying of Nucleic Acid Templates aus der Sammlung von iBiology
Kommentare und Kritik von Fachleuten, Lernenden und deren Eltern sind jederzeit willkommen.
Roland Heynkes, CC BY-NC-SA 4.0
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}