
Lerntext Blutzellen
Roland Heynkes 22.3.2025, CC BY-SA-4.0 DE
If you cannot read German-language texts, you could try a translation into your native language with DeepL.
Diese Internetseite soll erklären, was man wissen muss, um unsere Blutzellen einigermaßen zu verstehen.
| Erklärung unbekannterer Fachbegriffe |
|---|
| Hämopoese |
| Blutzellen |
| Die Vererbung der Blutgruppen A, B und 0 |
| Quellen |
|
Wir haben viele unterschiedliche Blutzelltypen, die aber alle von einer Sorte von Blutstammzellen abstammen. Das folgende Schema zeigt im Überblick die Hämopoese genannte Differenzierung von den Blutstammzellen zu den fertigen, spezialisierten Blutzellen. Man sieht im Überblick die Orte und Zellen des hämopoetischen Systems. Selbstverständlich habe ich auch darauf geachtet, die verschiedenen Zelltypen ungefähr im gleichen Maßstab darzustellen. Die Quellennachweise für die Bilder findet man im Glossar.
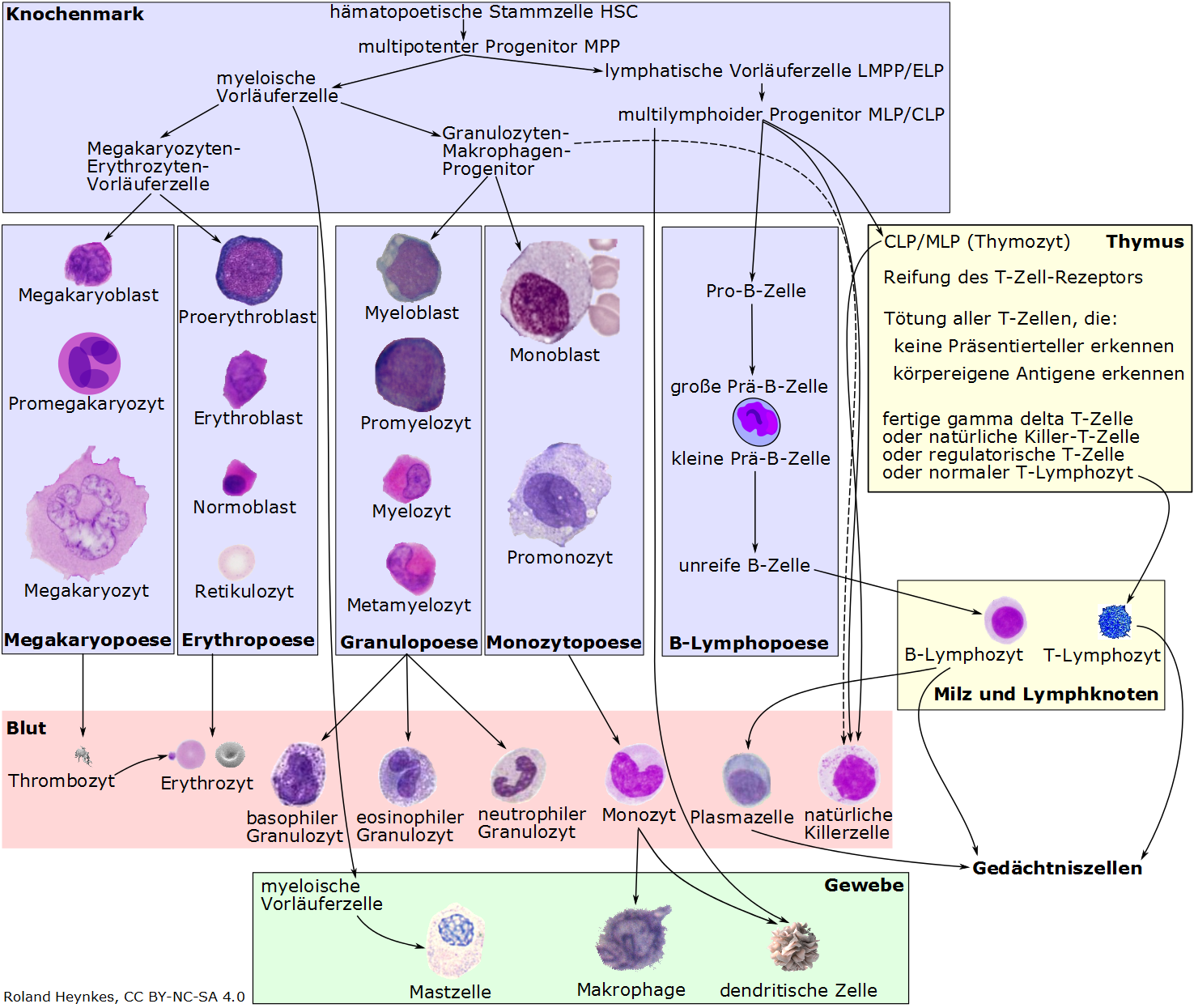
Nach einer aktuellen Schätzung besteht ein 20-30 Jahre alter, 70 kg schwerer und 170 cm großer "Referenzmann" aus durchschnittlich ungefähr 30 Billionen (30x1012) menschlichen Zellen und grob geschätzt 38 Billionen Bakterien (38x1012). Von den menschlichen Zellen stellen die roten Blutkörperchen mit rund 25 Billionen etwa 84%. Die nur rund 5 Liter Blut im Körper eines durchschnittlichen Erwachsenen können nur deshalb etwa 84% aller menschlichen Zellen enthalten, weil unsere roten Blutkörperchen (Erythrozyten) so klein sind. Streng genommen sind rote Blutkörperchen gar keine lebendigen Zellen mehr, denn sie besitzen keinen Zellkern und damit keinen Bauplan mehr. Im Gegensatz zu unseren anderen Körperzellen produzieren Erythrozyten deshalb auch keine MHC-1-Präsentierteller.
Unsere Erythrozyten sind einem großen Abnutzungsstress ausgesetzt, denn sie werden täglich schätzungsweise anderthalb Tausend mal durch den Lungen- und anschließend durch den Körperkreislauf mit ihren engen Kapillaren gepresst. Da sie keinen Zellkern mehr besitzen, können sie sich nicht reparieren. Die Dokumentation "Der menschliche Körper" nennt als durchschnittliche "Lebensdauer" nur 3 Monate, nach anderen Quellen sind es 4 Monate. Bei einer angenommenen "Lebensdauer" von nur 3 Monaten müsste ein 70-kg-Mann jeden Monat 25/3=8,33 Billionen, jeden Tag etwa 274 Milliarden, pro Stunde gut 11,4 Milliarden, pro Minute rund 190 Millionen und pro Sekunde fast 3,2 Millionen neue rote Blutkörperchen produzieren. Wenn die Erythrozyten 4 Monate durchhalten, dann müssen immer noch fast 2,4 Millionen rote Blutkörperchen pro Sekunde ersetzt werden.
Unsere noch lebenden Körperzellen produzieren noch ständig neue MHC-1-Präsentierteller. Auf ihnen präsentieren sie Bruchstücke (Peptide) von allen Eiweiß-Sorten, die sie herstellen. T-Killerzellen kontrollieren die auf MHC-1-Präsentiertellern befindlichen Peptide. Passen die Rezeptoren einer T-Killerzelle auf ein so präsentiertes Peptid, dann leitet die T-Killerzelle die Selbstzerstörung der betroffenen Zelle ein. Weil T-Killerzellen normalerweise nur körperfremde Peptide erkennen, ist eine Zelle mit fremden körperfremden Peptiden auf ihren MHC-1-Präsentiertellern sehr wahrscheinlich eine Krebszelle oder von Viren befallen. Wenn allerdings der Rezeptor einer T-Killerzelle doch ein körpereigenes Peptid erkennt, dann kann sie die Ausrottung dieses Zelltyps im ganzen Körper einleiten und damit eine Autoimmunkrankheit wie die Multiple Sklerose oder Typ-1-Diabetes auslösen. Oft ist eine Infektionskrankheit Auslöser einer Autoimmunkrankheit. Die eigentliche Ursache ist aber, dass eine T-Killerzelle im Thymus nicht getötet wurde, obwohl ihr Rezeptor ein körpereigenes Peptid erkannte.
MHC-2-Präsentierteller werden nur von speziellen Antigen-präsentierenden Zellen benutzt, um die Antigene von Krankheitserregern zu präsentieren, die sie gefressen haben. Das tun B-Zellen und verschiedene Arten von Fresszellen. MHC-2-Präsentierteller werden von T-Helferzellen kontrolliert. Passen ihre Rezeptoren auf ein darauf präsentiertes Peptid, dann wird die T-Helferzelle aktiviert, vermehrt sich und ihre Tochterzellen machen sich auf die Suche nach ihren Zielzellen.
Der genaue Funktions-Mechanismus ist noch nicht bekannt, aber häufig lassen sich Allergien durch eine sogenannte Hyposensibilisierung lindern.
Bestimmte Stoffe können bei manchen Menschen bei Berührung eine sogenannte Kontaktallergie auslösen.
B-Zellen sind noch nicht aktivierte Lymphozyten, die in geringen Mengen Antikörper produzieren und auf ihren Oberflächen präsentieren. Stößt zufällig ein Krankheitserreger mit einem passenden Antigen auf solche einen Antikörper, dann kann er gebunden werden. Dann wird der Krankheitserreger mit Hilfe des oder mehrerer gebundener Antikörper in die Zelle gezogen und darin zerlegt. An die Antikörper gebundene Antigene werden dann an MHC-2-Präsentierteller gebunden und auf der Zelloberfläche präsentiert.
Für Impfungen werden Krankheitserreger wie Viren, Bakterien oder einzellige Eukaryoten durch Bestrahlung abgeschwächt oder durch Hitze zerlegt. Die Gentechnik macht es auch möglich, nur bestimmte Proteine eines Krankheitserregers und damit besonders sichere Impfstoffe zu produzieren.
Fresszellen fressen die Impfstoffe. Wenn nötig zerlegen sie diese. Dann präsentieren die Fresszellen die Antigene des Impfstoffs auf ihren MHC-2-Präsentiertellern. Dort werden sie von T-Helferzellen abgetastet.
T-Helferzellen wandern von Fresszelle zu Fresszelle und kontrollieren, ob auf deren MHC-2-Präsentiertellern Peptide liegen, die an die Rezeptoren der T-Helferzellen binden können. Passen ihre Rezeptoren auf ein darauf präsentiertes Peptid, dann wird die T-Helferzelle aktiviert, vermehrt sich durch Zellteilung und ihre Tochterzellen machen sich auf die Suche nach ihren Zielzellen. Die richtige Zielzelle ist eine B-Zelle, auf deren MHC-2-Präsentiertellern Peptide liegen, die auch die Rezeptor der T-Helferzelle binden kann. Hat die T-Helferzelle solch eine B-Zelle gefunden, dann wird die B-Zelle aktiviert, vermehrt sich und verwandelt sich (differenziert) dabei zu einer Plasmazelle, die massenhaft den genau auf den Krankheitserreger passenden Antikörper produziert.
Ein Teil unserer Lymphozyten reift im Thymus. Deshalb nennt man sie T-Lymphozyten. Die Reifung der T-Lymphozyten bedeutet, dass sie aus einer größeren Auswahl von Bauplan-Bruchstücken nach dem Zufallsprinzip eine Bauanleitung (Gen) für einen T-Zell-Rezeptor erzeugen. So bekommt jeder T-Lymphozyt ein einmaliges Gen für einen einzigartigen T-Zell-Rezeptor. Deshalb kann jeder T-Zell-Rezeptor ein anderes Antigen binden. Gleichzeitig kann jeder T-Zell-Rezeptor nur ein Antigen binden. Das Problem bei der T-Zellreifung ist, dass die nach dem Zufallsprinzip erzeugten T-Zell-Rezeptoren häufig Antigene des eigenen Körpers erkennen. Wenn solche T-Lymphozyten lebend aus dem Thymus entkommen, können sie eine Autoimmunkrankheit verursachen. Deshalb werden alle gerade gereiften T-Lymphozyten im Thymus einem zweifachen Test unterzogen. Bestehen sie die Tests nicht, müssen sie sterben.
Der eine Test betrifft die Eignung des Rezeptors, Präsentierteller zu erkennen. Erkennt er MHC-1-Präsentierteller, dann wird der T-Lymphozyt eine T-Killerzelle. Erkennt er MHC-2-Präsentierteller, dann wird der T-Lymphozyt eine T-Helferzelle. Erkennt er aber gar keine Präsentierteller, dann ist er nutzlos und wird vernichtet.
Der andere Test ist der entscheidende. Ob ein Rezeptor körperfremde Antigene erkennt, lässt sich nicht testen. Aber wenn der Rezeptor eines T-Lymphozyten ein körpereigenes Antigen erkennt, dann muss er sterben.
Die Vererbung der Blutgruppen wird im Lerntext: "Vererbung der Blutgruppen A, B und 0" erklärt.
Sehr gute Mikroskop-Fotos von verschiedenen Blutzellen findet man hier (Use these images to learn more about your Life's Blood, or to enliven science or health papers with accurate images).

