
Lerntext Zellbiologie
Roland Heynkes 22.3.2025, CC BY-SA-4.0 DE
If you cannot read German-language texts, you could try a translation into your native language with DeepL.
Dieser Schulbuch-unabhängige Selbstlern-Hypertext soll es Schülern (m,w,d) ermöglichen, selbständig und im eigenen Tempo zu lernen oder zu wiederholen, was gebildete Menschen über Zellbiologie wissen sollten und was in der Qualiphase der gymnasialen Oberstufe als Vorwissen erwartet wird. Zur Vermeidung überflüssiger und für die Weiterentwicklung meiner Lerntexte hinderlicher Redundanzen habe ich einige Kapitel dieses Selbstlern-Hypertextes in passende andere buchunabhängige Lerntexte ausgelagert.
Menschen können nur leben, weil uns unsere Haut von unserer Umwelt trennt, während im Inneren unsere Organe genau aufeinander abgestimmt zusammen arbeiten. In unserer Haut und unseren übrigen Organen leben extrem kleine Zellen als kleinste lebensfähige Einheiten und Grundbausteine unserer Körper. Und ähnlich wie der ganze menschliche Organismus besitzen auch unsere Zellen eine schützende Hülle und im Inneren winzige Organellen mit unterschiedlichen Aufgaben. Bei tierischen Zellen besteht die Hülle nur aus einer dünnen Zellmembran, die kontrolliert, was in die Zelle hinein und hinaus darf. Pflanzliche Zellen werden zusätzlich durch eine stabile Zellwand vor dem Platzen bewahrt. Tierischen und pflanzliche Zellen besitzen einen Zellkern mit dem Bauplan der Zelle. Er enthält Tausende Rezepte für die Herstellung der Proteine, die eine Zelle braucht. Außer der im Lichtmikroskop erkennbaren Zellwand unterscheiden sich pflanzliche von tierischen Zellen vor allem durch ihre grünen Chloroplasten. Diese können die Energie des sichtbaren Lichts nutzen, um aus Wasser und CO2 die energiereiche Glucose und nebenbei Sauerstoff zu produzieren.
Nicht nur ich erstelle Selbstlernmaterial wie diesen Lerntext. Andere können das zumindest technisch viel besser als ich. Deren Animationen und Lernvideos sind zwar nicht so genau auf meine Klassen und Kurse zugeschnitten, aber besonders für Interessierte bieten sie wunderbare Möglichkeiten für vertiefendes Selbststudium. Die folgenden Links sind Beispiele dafür zum Thema Zellbiologie.
Das Molecular and Cellular Biology Education Project des World Wide Web Instructional Committee der North Dakota State University stellt für PC und iPad kostenlos seine Virtual Cell Animation Collection allen Interessierten zur Verfügung.
Texte, Videos, Animationen und Simulationen zu einem breiten Sepktrum naturwissenschaftlicher Themen bietet auch das kostenlose SpongeLab.
Der WDR bietet eine Internetseite an, auf der man zum Ansehen und Nachlesen in einem Archiv nach einer ganzen Reihe auch für Schülerinnen und Schüler gut geeigneter Fernseh-Dokumentationen suchen kann. Hier sind die Ergebnisse (3 Seiten) einer Suche mit dem Suchwort Zelle.
Julie A. Theriot ist Professorin an der Stanford University School of Medicine. Von ihr findet man Vorträge vor allem über das Zytoskelett und die Bewegungsapparate in Zellen bei YouTube und iBiology.
Bei YouTube und iBiology findet man mehrere Vorträge über das Zytoskelett und Motorproteine von Prof. Ronald David Vale.
Von Prof. Satyajit Mayor findet man bei YouTube und iBiology mehrere Vorträge über Membrane Rafts.
Prof. Mina Bissell erforscht die Bedeutung der extrazellulären Matrix von Zellen für deren Differenzierung und Entartung. Man findet einige ihrer Vorträge bei YouTube und iBiology
Die unten aufgelisteten Einzel-Videos und anderes Selbstlernmaterial habe ich weder produziert noch ins Internet gestellt noch übernehme ich dafür irgendeine Verantwortung. Es liegt insbesondere in der Verantwortung von Youtube, sicherzustellen, das die von Youtube bereitgestellten Videos keine Raubkopien sind. So wurde unter anderem ein anscheinend nur für Käufer des Biologie-Lehrbuches: "Biology" legal verfügbares Video (Protein Synthesis Animation) von Youtube gelöscht. Weil also Youtube und die Urheber offenbar die für Internet-Nutzer gar nicht machbare Überprüfung der Legalität übernehmen, kann es nach meinem Rechtsverständnis nicht verboten sein, sich mit Hilfe von Youtube Videos anzusehen, die das Verständnis dessen erleichtern, was im Biologie-Unterricht gelernt werden soll. Leider muss man teilweise erst einen Werbevorfilm ertragen, bevor man das eigentliche Video sehen kann.
Youtube macht Lernenden viele interessante Lernvideos zu biologischen Themen zugänglich. Viele auf Deutsch:
Sehr viel mehr Biologie-Videos gibt es auf Englisch:
XVIVO Scientific Animation translates complex science into compelling medical media:
Der Film: "Die Zelle - Baustein des Lebens" zeigt, wie die Entwicklung von Mikroskopen und Färbetechniken biologisches Wissen erweiterte. Letztlich entwickelten sich daraus die Zellbiologie und die Mikrobiologie. Man kann sich diesen kurzen Film seit Jahren in zahlreichen Versionen bei YouTube ansehen. Ich habe ihn in einem eigenen Lerntext ausführlich zusammengefasst. Wir übten mit ihm auch das kritische Mitdenken und die Suche nach Gegenbeispielen zur Widerlegung im Film gemachter Behauptungen. Wer sich intensiver mit der Entwicklung der Mikroskopie beschäftigen möchte, findet mehr Informationen in meinem kurzen Lerntext über das Mikroskop.
Die Biologie ist eine Naturwissenschaft, die früher hauptsächlich durch die Beobachtung natürlicher Phänomene gelernt hat. Immer wieder haben deshalb neue technische Möglichkeiten zu bedeutenden neuen Erkenntnissen in der Biologie geführt. Ein solcher Meilenstein war die Erfindung des Lichtmikroskops um das Jahr 1600, die 1665 zur Entdeckung der Zellen durch Robert Hooke und etwa 1675 zur Entdeckung von Einzellern und sogar Bakterien durch Antoni van Leeuwenhoek führte. Erst 1838 erkannten der Botaniker Matthias Schleiden und der forschende Mediziner Theodor Schwann aufgrund mikroskopischer Untersuchungen, dass alle Pflanzen und Tiere aus Zellen aufgebaut sind. Ebenfalls mit Hilfe eines Lichtmikroskops beobachtete Hugo von Mohl 1835 erstmals eine Zellteilung. Rudolf Virchow leitete aus dieser und vielen weiteren Beobachtungen 1855 die Hypothese ab, dass Zellen nur aus Zellen (durch Zellteilung) und nicht etwa aus Schlamm oder anderen nicht lebenden Materialien entstehen (Omnis cellula e cellula). Außerdem erkannte dieser berühmte Arzt, dass viele Krankheiten mit bestimmten Veränderungen von Zellen verbunden sind. Allerdings muss natürlich irgendwann eine allererste Zelle entstanden sein.
Bereits 1838 benutzte Christian Gottfried Ehrenberg den aus Schildläusen gewonnenen roten Farbstoff Karmin zur Anfärbung von Einzellern. Großen Anteil an der Entwicklung der Färbetechniken für Zellbestandteile hatte um das Jahr 1855 der Anatom Joseph von Gerlach. Mit einer eher zufälligen Entdeckung durch William Henry Perkin begann 1856 die Entwicklung künstlicher Textilfarbstoffe, die es Paul Ehrlich ermöglichten, 1879-1894 die Anfärbung von Zellen zu perfektionieren. Diese Entwicklung der Anfärbung von Zellen war wichtig, weil die unterschiedliche Anfärbbarkeit verschiedener Zellbestandteile diese unterscheidbar machte und Kontraste verstärkte.
Noch wichtiger als die Färbung ist für das Mikroskopieren die Fixierung der Zellbestandteile, damit sie nicht zerfallen, enzymatisch zersetzt oder später von Bakterien oder Pilzen gefressen werden. Bei der Untersuchung seiner desinfizierenden Eigenschaften entdeckte Ferdinand Blum 1893, dass sich Formalin (35-40%ige Formaldehydlösung) besser als Alkohol zur Fixierung und Konservierung mikroskopischer Präparate eignet. Diese Methode ist bis heute die gebräuchlichste.
Sehr wichtig für das Verständnis der Zelle war auch die Erfindung des Elektronenmikroskops 1931 durch Ernst Ruska und Max Knoll sowie etwa zeitgleich durch Reinhold Rüdenberg, weil dadurch die Feinstruktur der Zelle und damit das genaue Aussehen der Organellen sichtbar wurde. Auch die Viren wurden erst durch Elektronenmikroskope sichtbar.
Sehr anschaulich verständlich macht das Innere einer Zelle eine Animation des Abwehrkampfes einer Zelle gegen Adenoviren, die sich in ihr zu vermehren versuchen. Man kann diese Dokumentation manchmal im Fernsehen, aber auch jederzeit hier und hier im World Wide Web sehen.
Damit sich alle Lernenden voll auf die einmaligen Bilder konzentrieren können, habe ich eine kritische Zusammenfassung der Fernsehdokumentation: "evolutionärer Wettkampf Zelle gegen Viren" erstellt.
In diesem Films erkennt man wichtige Prinzipien der Biologie:
Informationen über die Organellen eukaryotischer Zellen findet man im Lerntext Organellen.
Den Aufbau einer tierischen Zelle findet man im Lerntext Organellen.
Jedes Lebewesen grenzt sich durch eine Hülle von seiner Umwelt ab. Das tut auch jede einzelne Zelle eines vielzelligen Eukaryoten mit einer extrem dünnen Membran aus Lipiden und Proteinen. Man nennt sie Zellmembran und sie ist semipermeabel. Der Wortteil "semi" bedeutet halb und der Wortteil "permeabel" bedeutet durchlässig. Die Zellmembran lässt selektiv nur bestimmte Dinge in die Zelle hinein oder aus ihr hinaus. Ihre Funktion entspricht der Aufgabe einer antiken oder mittelalterlichen Stadtmauer mit bewachten Stadttoren.
Die Membranen der Zellen bestehen zu großen Teilen aus Lipiden. Näheres zu den Lipiden findet man im Lerntext Biomoleküle. Die unpolaren Anteile der Lipide können keine Bindungen zu Wasser-Molekülen aufbauen und heißen deshalb hydrophob. Das folgende Schema der Zellmembran zeigt im Inneren der Membran gelb die Cholesterin-Moleküle und hellbraun die Fettsäuren. Diese unpolaren Anteile der Lipide werden durch schwache Van-der-Waals-Wechselwirkung zusammengehalten. So bilden sie die Grundsubstanz (Matrix) jeder Biomembran. Nur an einer Seite besitzen die Lipide als orange Kugeln dargestellte polare Anteile, mit denen sie Wasserstoffbrückenbindungen zu Wasser-Molekülen aufbauen können und die deshalb hydrophil heißen. Am anderen Ende sind die Lipide hydrophob und meiden den Kontakt zu Wasser. Darum bilden sie Doppelmembranen mit zwei hydrophilen Außenseiten und einem hydrophoben Inneren.
| Schema einer Zellmembran |
|---|
 |
| Mariana Ruiz Villarreal, public domain |
| Auf einer eigenen Seite kann man sich mittels dreier selbst manipulierbarer, dreidimensionaler JSmol-Darstellungen die Strukturen von Cholesterin, Palmitinsäure und der Omega-3-Fettsäure Docosahexaensäure von allen Seiten ansehen und diese Lipid-Moleküle dadurch besser verstehen. |
Das Schema zeigt, wie unterschiedlich Proteine mit einer Membran verbunden oder in sie integriert sein können. Auf der Zelloberfläche sieht man unterschiedliche Kohlenhydrate an den Proteinen und gebunden an Lipide. Die Kohlenhydrate ermöglichen unter anderem die Unterscheidung verschiedener Zelltypen.
Informationen zum Thema Transmembran-Transport findet man im Lerntext Transmembran-Transport.
Die Themen Zellkerne (Nukleus) eukaryotischer Zellen, Nukleoli, Zellkernhülle und Kernporen erarbeiteten wir uns mit dem Lerntext Organellen:
Ribosomen sind komplexe Strukturen, die sich im Cytoplasma aus zwei Untereinheiten zusammensetzen, wenn die kleinere Untereinheit eine mRNA gebunden hat. Die Untereinheiten entstehen in Nucleoli und gelangen durch Kernporen aus dem Zellkern ins Zytoplasma. Die Ribosomen-Untereinheiten sind keine Moleküle, sondern Komplexe aus rRNAs und Proteinen.
Ribosomen übersetzen in allen bekannten Lebewesen im Zytoplasma das selbe 4-Buchstabenalphabet der Nukleinsäuren in das 20-Aminosäuren-Alphabet der Proteine. Im Prozess der Translation liest ein Ribosom eine aus dem Zellkern ins Zytoplasma transportierte Boten-RNA (mRNA) genannte Kopie einer in DNA codierten Erbinformation (Gen). Aus der Reihenfolge (Sequenz) der Nukleotide in der mRNA ergibt sich die Sequenz der Aminosäuren eines Proteins. Dazu ist es erforderlich, dass jede einzelne Aminosäure an eine spezielle transfer-RNA (tRNA) gebunden ist. Am Ribosom binden direkt nebeneinander zwei tRNAs mit passenden Anticodons an benachbarte Codons einer mRNA und das Ribosom verbindet dann die an den tRNAs hängenden Aminosäuren. Die Ribosomen entsprechen also den Produktionsmaschinen oder -Robotern einer Stadt.
| Ribosomen-Untereinheiten | |
|---|---|
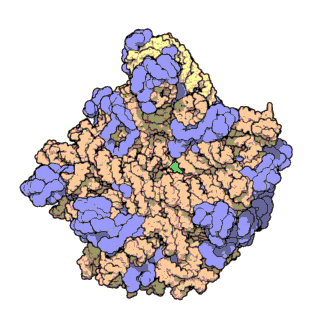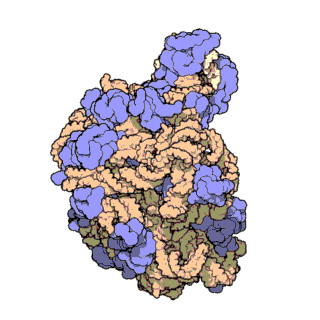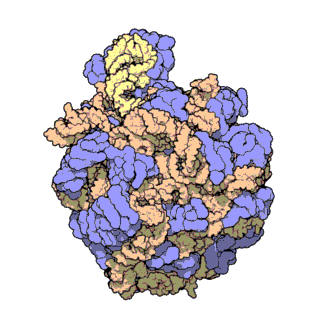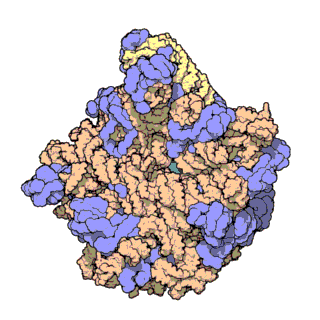 |
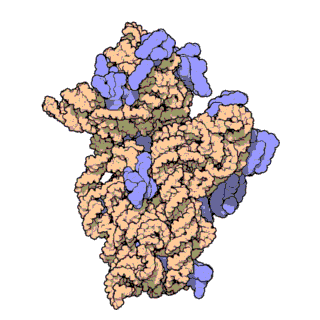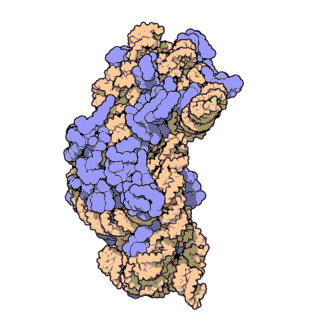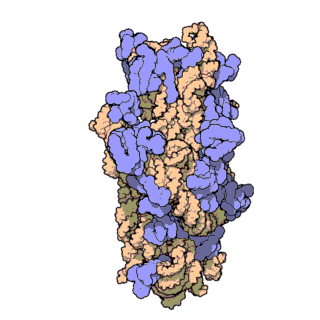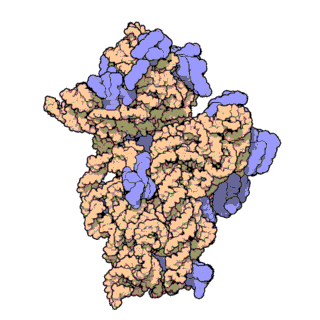 |
| Diese animierten GIF zeigen links die große und rechts die kleine Ribosomen-Untereinheit. | |
| Protein Data Bank Educational Resources | |
An eine mRNA können mehrere Ribosomen gleichzeitig binden. Man nennt solche Komplexe Polysomen oder Polyribosomen.
Die Ribosomen von Prokaryoten und Eukaryoten unterscheiden sich hinsichtlich ihrer Sedimentationsgeschwindigkeiten, die von der Größe und der Dichte abhängen und mit einer Zentrifugation gemessen werden. Tiere und Pflanzen haben größere Ribosomen als Bakterien, aber die Ribosomen von Chloroplasten und Mitochondrien entsprechen eher denen der Bakterien. Dies unterstützt die Endosymbionten-Theorie, nach welcher die Plastiden und Mitochondrien ursprünglich Bakterien waren, die heute in enger Endosymbiose mit Eukaryoten-Zellen leben.
Vesikel ist das lateinische Wort für Bläschen. Mediziner bezeichnen viele Strukturen des menschlichen Körpers als Vesikel. Zellbiologen meinen damit kleine Bläschen mit einer Hülle aus einer Membran, die als mikroskopisch kleine Reaktions- und Transporträume innerhalb eukaryotischer Zellen fungieren. Sie sind alle kugelig, haben aber unterschiedliche Funktionen und dementsprechend verschiedene Namen wie Endosomen, Lysosomen, Mikrokörperchen, Peroxisomen, Mikrosomen und Glyoxysomen. Unter Vesikeln im engeren Sinne versteht man die reinen Transportvesikel, auf die keine der oben genannten Bezeichnungen zutrifft. Diese Transport-Vesikel sind die Lastwagen oder Eisenbahnwagons einer Zelle. Vesikel entstehen durch Abschnürung von einer größeren Membranfläche wie dem endoplasmatischen Retikulum, den Dictyosomen des Golgi-Apparates, einem Endosom oder der Zellmembran. Über das Cytoskelett laufende Motorproteine ziehen Vesikel durch das Cytoplasma zu ihren Zielmembranen. Dort verschmelzen (fusionieren) sie mit der Zielmembran und entleeren dabei ihr Inneres auf die andere Seite der Zielmembran, also aus der Zelle heraus oder in Organellen hinein. Auf diese Weise sorgen Vesikel auch für einen ständigen Austausch von Membranen.
Informationen zum rauen endoplasmatischen Retikulum der eukaryotischen Zellen findet man im Lerntext Organellen.
Informationen zum glatten endoplasmatischen Retikulum der eukaryotischen Zellen findet man im Lerntext Organellen.
Informationen zum Golgi-Apparat der eukaryotischen Zellen findet man im Lerntext Organellen.
Informationen zu Membranfluss und Fließgleichgewichten in aktiv lebenden eukaryotischen Zellen findet man im Lerntext Membranfluss:
Mikrotubuli, Intermediärfilamente und Mikrofilamente bilden die Hauptmasse des Zytoskeletts eukaryotischer Zellen, welches der Zelle eine Form gibt, ihr Bewegung ermöglicht und als dreidimensionales Transportwege-Netzwerk genutzt wird.
Intermediärfilamente sind das Zytoplasma einer Zelle durchziehende Filamente aus Protein-Untereinheiten. Sie sind am Aufbau des Zytoskeletts beteiligt. Das "Inter" im Namen deutet an, dass Intermediärfilamente mit einer Dicke von etwa 10 Nanometern zwischen den nur 6-7 nm dicken Mikrofilamenten und den rund 25 nm dicken Mikrotubuli stehen. Intermediärfilamente helfen auch, Zellen aneinander zu binden. Mikrofilamente sind fadenförmige Protein-Strukturen mit nur 6 Nanometern Durchmesser in eukaryotischen Zellen. Sie bestehen hauptsächlich aus dem Protein Aktin und werden daher auch als Aktin-Filamente bezeichnet. Mikrotubulus nennen Biologen ein röhrenförmiges, 20-30 nm dickes Filament aus den Proteinen α-Tubulin und β-Tubulin.
Informationen zu den Lysosomen der eukaryotischen Zellen findet man im Lerntext Organellen.
Das nur scheinbar strukturlose Cytoplasma ist die viskose, leicht körnige Grundsubstanz der Zellen, in der Ribosomen, Filamente sowie die Organellen schwimmen und eine Kompartimentierung schaffen. Es besteht zu 60-90% aus Wasser, in dem anorganische Moleküle und Ionen gelöst sind, aber auch organische Moleküle, darunter vor allem Proteine, Lipide, Polysaccharide und RNA. Seine Konsistenz kann zwischen einem flüssigen Sol- und einem festeren Gelzustand wechseln. In ihm finden unzählige Stoffwechselreaktionen statt.
Informationen zu den Peroxisomen eukaryotischer Zellen findet man im Lerntext Organellen.
Zentriol oder Centriol nennen Zellbiologen eine meist paarweise auftretende, zylinderförmige Struktur mit einer Größe von etwa 170 x 500 Nanometern. Man findet sie in den meisten tierischen sowie den Zellen niederer Pflanzen. Zentriole bilden zusammen mit der perizentriolaren Protein-Matrix das Zentrosom und organisieren die Mikrotubuli. In begeißelten Zellen liegen die Zentriolen in der Interphase als geißeltragende Basalkörper vor. Die Zentriolen der meisten Säugetiere bestehen aus 27 Mikrotubuli (9 Tripletts), die über dünnere Filamente miteinander verbunden sind. Bei der Fruchtfliege Drosophila bestehen die Zentriolen aus Mikrotubuli-Dupletts und beim Fadenwurm C. elegans sind es Mikrotubuli-Singletts.
| Schema eines Zentriols |
|---|
 |
| anonym, CC BY-SA 3.0 |
Zentrosom oder Centrosom bzw. Zentralkörperchen nennen Zellbiologen eine aus zwei einen 90-Grad-Winkel bildenden Zentriolen und einer diese umgebenden perizentriolaren Protein-Matrix bestehende Struktur, die in den meisten tierischen sowie den Zellen niederer Pflanzen, aber nicht in den modernen (Angiospermen) Pflanzen vorkommt. Wie der Name schon andeutet, befindet sich das Zentrosom in der Interphase zentral in der Nähe des Zellkerns. Je nach der bevorzugten Organell-Definition wird es zu den Organellen gezählt oder nicht. Zentrosome organisieren in der Interphase den mikrotubulären Teil des Zytoskeletts und während der Mitose und der Meiose den Spindelapparat zur Trennung der Chromatiden bzw. Chromosom und werden deshalb auch microtubule organizing center genannt. Wenn die DNA-Replikation aus 1-Chromatid-Chromosomen 2-Chromatiden-Chromosomen macht, dann verdoppelt sich auch das Centrosom und die beiden Centrosomen wandern aus der Zellmitte zu zwei Polen, von denen aus sie den Spindelapparat organisieren.
Informationen zu den Endosomen eukaryotischer Zellen findet man im Lerntext Organellen.
Eukaryotische Zellen enthalten zusätzliche Erbinformationen in Mitochondrien und bei pflanzlichen Zellen außerdem in Chloroplasten. Mitochondrien und Chloroplasten enthalten ihre eigenen kleinen Baupläne, weil sie wie winzige Bakterien in eukaryotischen Zellen leben und sich selbständig durch Zellteilung vermehren. Insofern ist schon jede einzelne eukaryotische Zelle ganz ähnlich wie eine Flechte eine Symbiose (genauer: Endosymbiose) aus einer großen schützenden Zelle und ihren lebenswichtige Energie liefernden Endosymbionten. Wegen dieser relativen Unabhängigkeit (Semiautonomie) der Mitochondrien werden ihre Baupläne oft vergessen, wenn von der Genom genannten Gesamtheit aller Erbinformationen eines eukaryotischen Lebewesens die Rede ist. Das ist aber nicht korrekt, sofern man beispielsweise uns Menschen als Lebewesen und nicht nur als Symbiose zwischen Mensch und Mitochondrien betrachtet.
Informationen zur Endosymbionten-Theorie findet man im Lerntext Endosymbionten-Theorie.
Informationen zu den Mitochondrien der eukaryotischen Zellen findet man im Lerntext Organellen.
Zu den folgenden Kapiteln gibt es ein Lernmodul mit Klausur-ähnlichen Aufgaben (pdf).
| Schema einer Pflanzenzelle | |
|---|---|
 | |
| Mariana Ruiz Villarreal, public domain | |
Pflanzenzellen sind eine Weiterentwicklung tierischer Zellen. Zusätzlich zu den Struktur-Elementen tierischer Zellen besitzen sie Plastiden für unterschiedliche Aufgaben. Wie die Mitochondrien sind auch diese Plastiden Endosymbionten, die sich dank eigener Baupläne selbständig vermehren können.
Plasmalemma nennt man bei Pflanzenzellen die Zellmembran, die das Cytoplasma einschließt und von der Umgebung der Zelle abgrenzt. Das Plasmalemma wird außen noch durch eine inneren Druck abfangende Zellwand begrenzt, weil Pflanzenzellen ohne Zellwand platzen würden.
Zwischen Pflanzenzellen gibt es ganz dünne Mittellamellen, die sich während der Zellteilung aus Golgi-Vesikeln bilden und nachher die Nachbarzellen miteinander verkleben. Und in Pflanzenzellen gibt es Vakuolen, die immer größer werden und miteinander verschmelzen, sodass am Ende eine riesige Vakuole die ganze Pflanzenzelle dominiert und alle Organellen an den Rand drängt. Die Vakuolen dienen als Speicher für Vorräte, Farbstoffe und Abfallstoffe. Einige dieser Stoffe können derart konzentriert sein, dass sie nicht mehr gelöst werden können und deshalb Kristalle bilden.
Prall mit Wasser gefüllte Bakterien und Pflanzenzellen würden platzen, hätten sie nicht eine zugfeste Zellwand, die dem inneren Wasserdruck standhält. Sie besteht aus einem Geflecht langkettiger und oft verzweigter Makromoleküle. Der wichtigste Baustoff der Zellwand ist die Zellulose. Cellulose ist ein unverzweigtes Polysaccharid aus 1.000 - 10.000 Glucose-Molekülen. Elementarfibrillen nennt man Bündel aus 50-100 Zellulosefäden. Innerhalb dieser Elementarfibrillen gibt es mehrere Micellen genannte Abschnitte, in denen die Cellulosefäden parallel verlaufen, während sie zwischen den Micellen durcheinander laufen und auf diese Weise ein Auseinanderfallen der Elementarfibrillen verhindern. Bündel von jeweils 15-20 Elementarfibrillen nennt man Mikrofibrillen. Das Zellulosegerüst kann in eine Grundsubstanz aus quellbaren Polysacchariden und einem geringeren Anteil von Proteinen eingebettet sein. Bei der Verholzung wird diese weichere Grundsubstanz durch die erheblich härtere, zug- und druckfestere Grundsubstanz Lignin ersetzt. Wenn eine noch wachsende Pflanzenzelle eine neue Zellwand bildet, dann werden die Mikrofibrillen in scheinbar regellosen Kurven angelegt. Man nennt das Streutextur und sie macht die Zellwand dehnungsfähig, weil die Kurven gerade gezogen werden können. Dadurch nimmt die Fläche der jungen Zellwand zu, während ihre Dicke abnimmt. Die Verdünnung der immer stärker gestreckten Primärwand wird ausgeglichen durch das Aufbringen immer neuer Fibrillennetze. Die Unterschiedlichkeit der Zellwandschichten kommt dadurch zustande, dass die Schichten umso weniger gedehnt werden, je später in der Wachstumsphase sie angelegt werden. Sie müssen deshalb nicht mehr als Streutextur angelegt werden, sondern werden gleich in Form parallel verlaufender Mikrofibrillen produziert, wobei die verschiedenen Schichten unterschiedlich ausgerichtet sind, um die Stabilität zu erhöhen.
Informationen zu den Plastiden eukaryotischer Zellen findet man im Lerntext Organellen.
Informationen zu den Chloroplasten eukaryotischer Pflanzenzellen findet man im Lerntext Organellen.
Informationen zu den Vakuolen eukaryotischer Pflanzenzellen findet man im Lerntext Organellen.
Im Lerntext Genetik findet man Informationen zu den Themen:
Im Lerntext Genetik findet man auch Informationen zu den Themen:
Wie die Monomere und Lipide aussehen, zeigt der Lerntext Biomoleküle.
Eine Tabelle mit Biopolymer-Fachbegriffen findet man im Lerntext Biomoleküle.
In den folgenden Kapiteln werden die Peptide und die Nukleinsäuren näher betrachtet.
Mit dem Lerntext Biomoleküle erarbeiteten wir uns die Themen:
Ebenfalls mit dem Lerntext Biomoleküle erarbeiteten wir uns die Themen:
Jedes Lebewesen besitzt wenigstens zeitweise mindestens eine Zelle. Jede noch teilungs- oder wenigstens noch wachstumsfähige Zelle benötigt einen Bauplan. Besitzt eine Zelle genau einen Bauplan und sollen bei einer Zellteilung beide Tochterzellen einen Bauplan erhalten, dann muss vor oder während der Zellteilung der Bauplan verdoppelt werden. Besteht der Bauplan aus doppelsträngiger DNA, dann werden zu diesem Zweck die beiden DNA-Stränge getrennt. Freie Nukleotide binden an passenden Basen beider DNA-Einzelstränge und werden zu komplementären Gegensträngen miteinander verbunden. Durch diesen im Prinzip einfachen Mechanismus entstehen aus einem DNA-Doppelstrang zwei identische Kopien. Das folgende, von Mariana Ruiz dankenswerterweise der Menschheit geschenkte Schema deutet trotz immer noch starker Vereinfachung an, dass die DNA-Replikation im Detail natürlich doch wieder nicht ganz so einfach ist. Man folge daher bei sämtlichen unbekannten Begriffen bitte den auch in den Beschriftungen funktionierenden Links in das HTML-Bio-Glossar.
| Schema der DNA-Replikation |
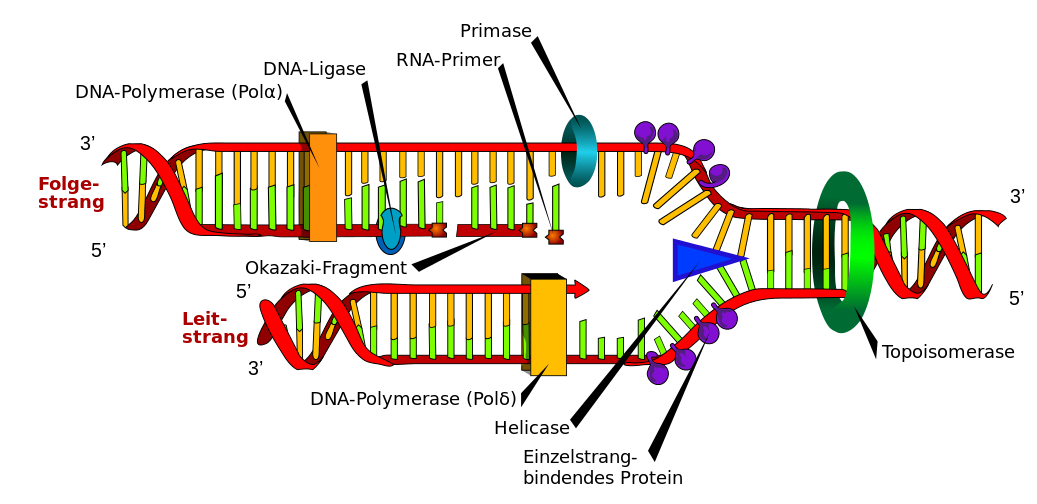
|
| Mariana Ruiz, gemeinfrei. Ich habe nur die Beschriftungen mit Koordinaten versehen, damit man mit einem Klick zum entsprechenden Eintrag im Glossar springen kann. |
An einer Replikationsgabel wie der oben gezeigten schafft die DNA-Replikation bei Bakterien (E. coli) ungefähr 500 bis 1000 Nukleotide pro Sekunde, in menschlichen Zellen sind es nur rund 50 Nukleotide pro Sekunde. Weil die DNA-Replikation in den Chromosomen von Eukaryoten an vielen Stellen beginnt, dauert die S-Phase trotzdem durchschnittlich nur rund 8 Stunden bei Säugetieren und 40 Minuten bei Hefe. (Quelle: Chapter 5, The Initiation and Completion of DNA Replication in Chromosomes in Alberts B, Johnson A, Lewis J, et al. - Molecular Biology of the Cell. 4th edition - New York: Garland Science; 2002.
Während der DNA-Replikation der 1-Chromatid-Chromosomen zu 2-Chromatiden-Chromosomen verdoppelt sich in der Nähe des Zellkerns auch das Centrosom mit seinen beiden Zentriolen und die beiden Centrosomen beginnen ihre Wanderung aus der Zellmitte zu zwei Zellpolen, von denen aus sie den Spindelapparat organisieren.
Nach der DNA-Replikation muss nur noch dafür gesorgt werden, dass jede Tochterzelle eine Kopie erhält. Bei sich alle 20 Minuten teilenden Bakterien werden allerdings die gerade als Ergebnis einer noch laufenden DNA-Replikation entstehenden Tochterchromosomen schon wieder einer weiteren Replikation unterzogen, damit die Vermehrung der Bakterien-Chromosomen mit der extremen Zellteilungsrate mithalten kann.
Bei Eukaryoten ist die Sache komplizierter. Im vollständigen Zellzyklus der Eukaryoten unterscheidet man meistens nicht nur zwischen den beiden Phasen Wachstum (Interphase) und Zellteilung. Sinnvoll ist zunächst innerhalb der Zellteilung eine Unterscheidung zwischen Kernteilung (Mitose) und Plasmateilung (Cytokinese), weil auf eine Kernteilung nicht immer eine Plasmateilung folgt. Die normale Mitose wird traditionell in Prophase, (Prometaphase), Metaphase, Anaphase und Telophase unterteilt, während man in der Wachstumsphase zwischen G1-, S- und G2-Phase unterscheidet. In der G1-Phase wächst eine Zelle ungefähr zur ursprünglichen Größe heran. Dann folgt die S-Phase, in der die DNA-Replikation, stattfindet. Ihr folgt die G2-Phase als Pause zwischen der Verdopplung des Bauplans und der Kernteilung. Es folgt die Kernteilung (Mitose) als Voraussetzung für die Plasmateilung (Cytokinese). Um den Zellzyklus abzuschließen, muss nun nur noch die (Cytokinese) erfolgen, die aus einer großen zwei kleine Zellen macht. Dazu werden in der Äquatorialebene zwei neue Zellmembranen und bei Pflanzen zusätzlich zwei Zellwände aufgebaut.
| Der Zellzyklus bei Eukaryoten | |
 |
|
Mit dem für meinen Geschmack entschieden zu teuren Programm: "Adobe After Effect" hat Jessica Scheiderer im Rahmen ihrer Bachelor-Arbeit ein tolles Video zur Erklärung der Mitose im Zellzyklus produziert.
Mitose oder Kernteilung nennen Biologen den Teil des Zellzyklusses, der von der Prophase bis zur vollendeten Telophase reicht.
Wer die ebenso eintönige wie überflüssige Hintergrundmusik erträgt, kann sich die Mitose durch ein von Jessica Scheiderer im Rahmen ihrer Bachelor-Arbeit produziertes Video zur Erklärung der Mitose im Zellzyklus erklären lassen.
Die bereits während der DNA-Replikation verdoppelten Centrosomen wandern während der Prophase weiter zu den beiden Zellpolen. Zwischen den beiden Centrosomen entwickelt sich der Spindelapparat.
In der Prophase kondensieren (ballen sich zusammen) die Chromosomen zu dicken, aber kurzen und damit transportfähigen Einheiten. In dieser Phase sieht man, dass jedes Chromosom aus zwei Schwesterchromatiden besteht. Das sind zwei nahezu identische Kopien des selben Chromosoms, die noch am Centromer aneinander gebunden sind und deshalb beide als Bestandteile eine 2-Chromatiden-Chromosoms bezeichnet werden.
| Schema der Prophase einer tierischen Zelle |

|
| anonymer Amerikaner, Roland Heynkes, CC BY-SA-4.0 |
In der Prometaphase wird die Kernhülle aufgelöst und die Mikrotubuli des Spindelapparates wachsen bis zu den Centromeren der Chromosomen.
| Schema der Prometaphase einer tierischen Zelle |

|
| anonymer Amerikaner, Roland Heynkes, CC BY-SA-4.0 |
In der Metaphase werden die Chromosomen vom Spindelapparat alle in eine Ebene in der Mitte der Zelle bugsiert, die man Äquatorialebene nennt.
| Schema der Metaphase einer tierischen Zelle |

|
| anonymer Amerikaner, Roland Heynkes, CC BY-SA-4.0 |
In der Anaphase zieht der Spindelapparat die beiden Schwesterchromatiden der Chromosomen auseinander bis an die gegenüber liegenden Pole der Zelle. Weit von einander entfernt liegt nun in beiden Hälften einer Zelle von jedem Chromosom jeweils ein Chromatid und man bezeichnet deshalb die Chromosomen von nun an bis zur nächsten S-Phase als 1-Chromatid-Chromosomen.
| Schema der Anaphase einer tierischen Zelle |

|
| anonymer Amerikaner, Roland Heynkes, CC BY-SA-4.0 |
In der Telophase löst sich der Spindelapparat auf, an den beiden Zellpolen bilden sich um die 1-Chromatid-Chromosomen herum neue Kernhüllen und in diesen dekondensieren die Chromosomen wieder. Das bedeutet, dass sie wieder ganz lang und dünn werden, sodass man keine einzelnen Chromosomen mehr erkennen kann. Man sieht nur noch eine einzige Masse, die aufgrund ihrer Färbkarkeit Chromatin nennt.
| Schema der Telophase einer tierischen Zelle |

|
| anonymer Amerikaner, Roland Heynkes, CC BY-SA-4.0 |
In Schemata sind die Stadien der Mitose relativ leicht zu erkennen. In echten, gefärbten Zellen gelingt das nicht so leicht. Die folgenden Fotos dienen der Übung.
| Die Metaphase einer Zelle (Mitte) in der Wurzelspitze einer Küchenzwiebel (Allium cepa) |
|---|
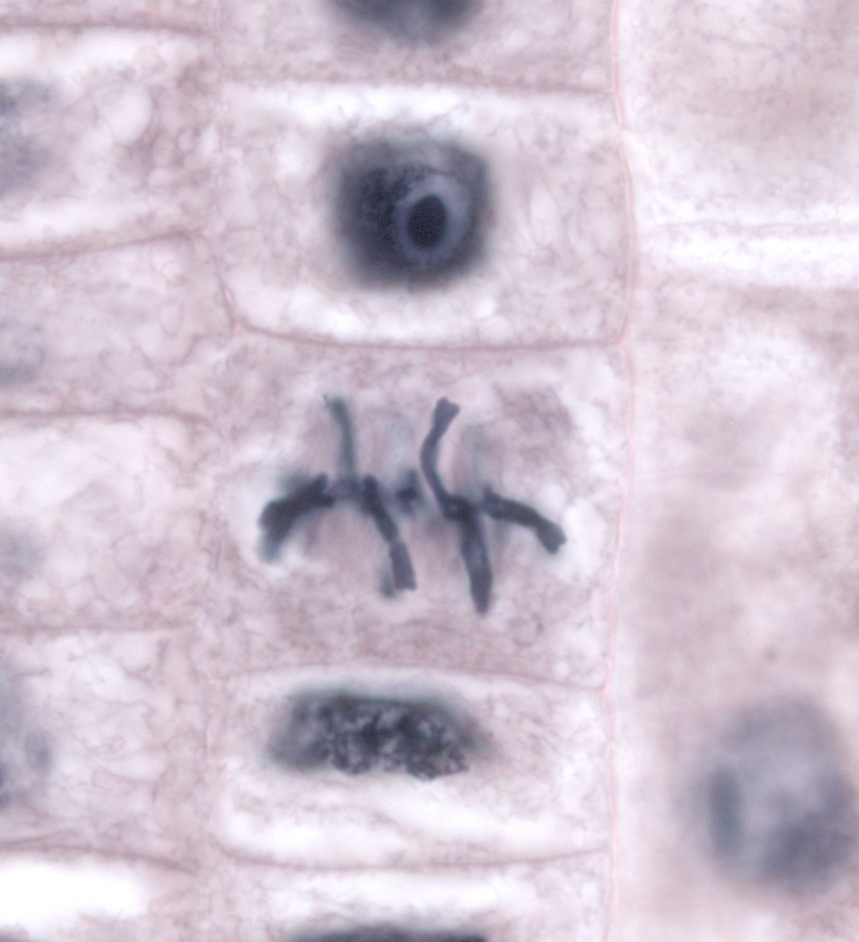
|
| Thomas Geier, CC BY-SA 3.0 |
|
Interphase oberhalb der Mitte, unterhalb der Mitte späte Metaphase oder frühe Anaphase in Wurzelzellen einer Küchenzwiebel (Allium cepa) |
|---|

|
| Thomas Geier, CC BY-SA 3.0 |
| Prophase (oben) und Anaphase (unten) in Wurzelzellen einer Küchenzwiebel (Allium cepa) |
|---|
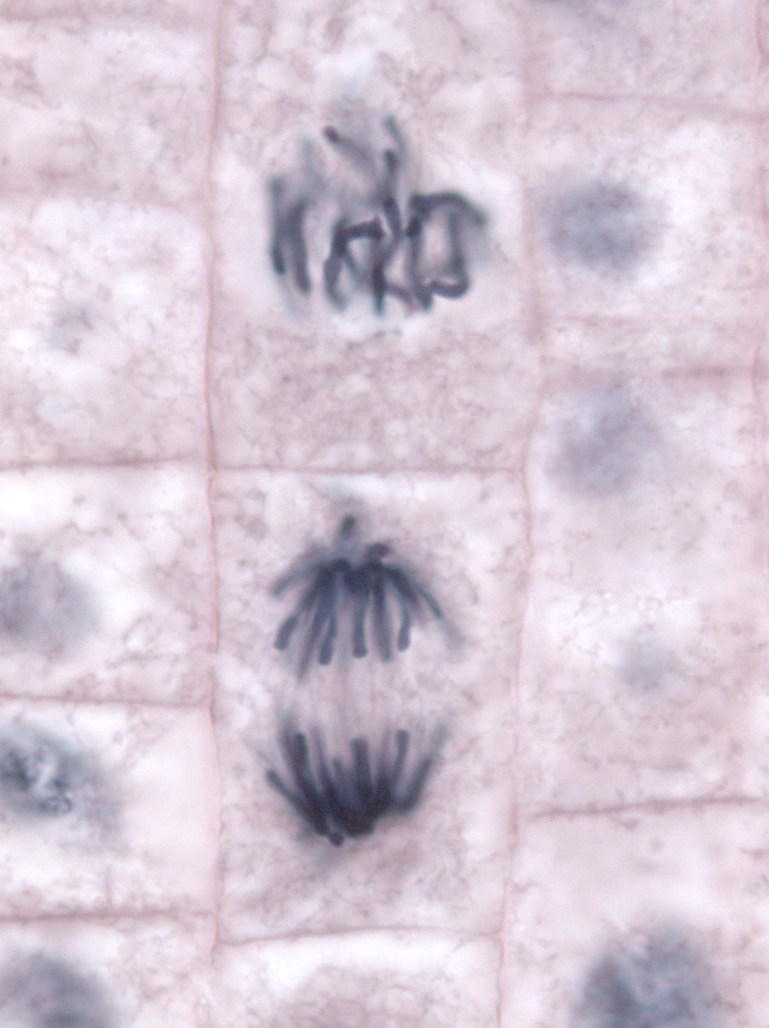
|
| Thomas Geier, CC BY-SA 3.0 |
| Telophase in Wurzelzellen einer Küchenzwiebel (Allium cepa) |
|---|
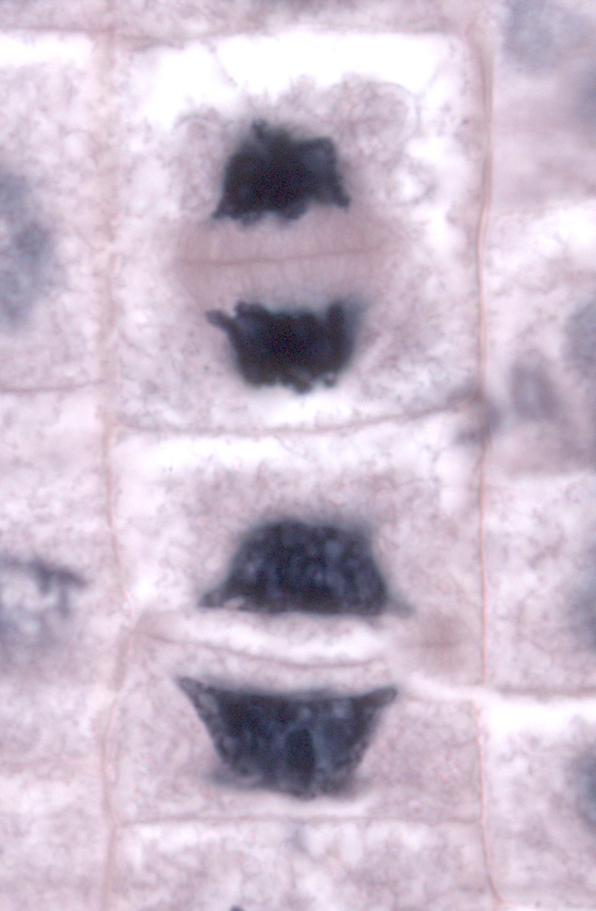
|
| Thomas Geier, CC BY-SA 3.0 |
| Zwiebelwurzelzellen in Inter-, Pro- und Anaphase |
|---|
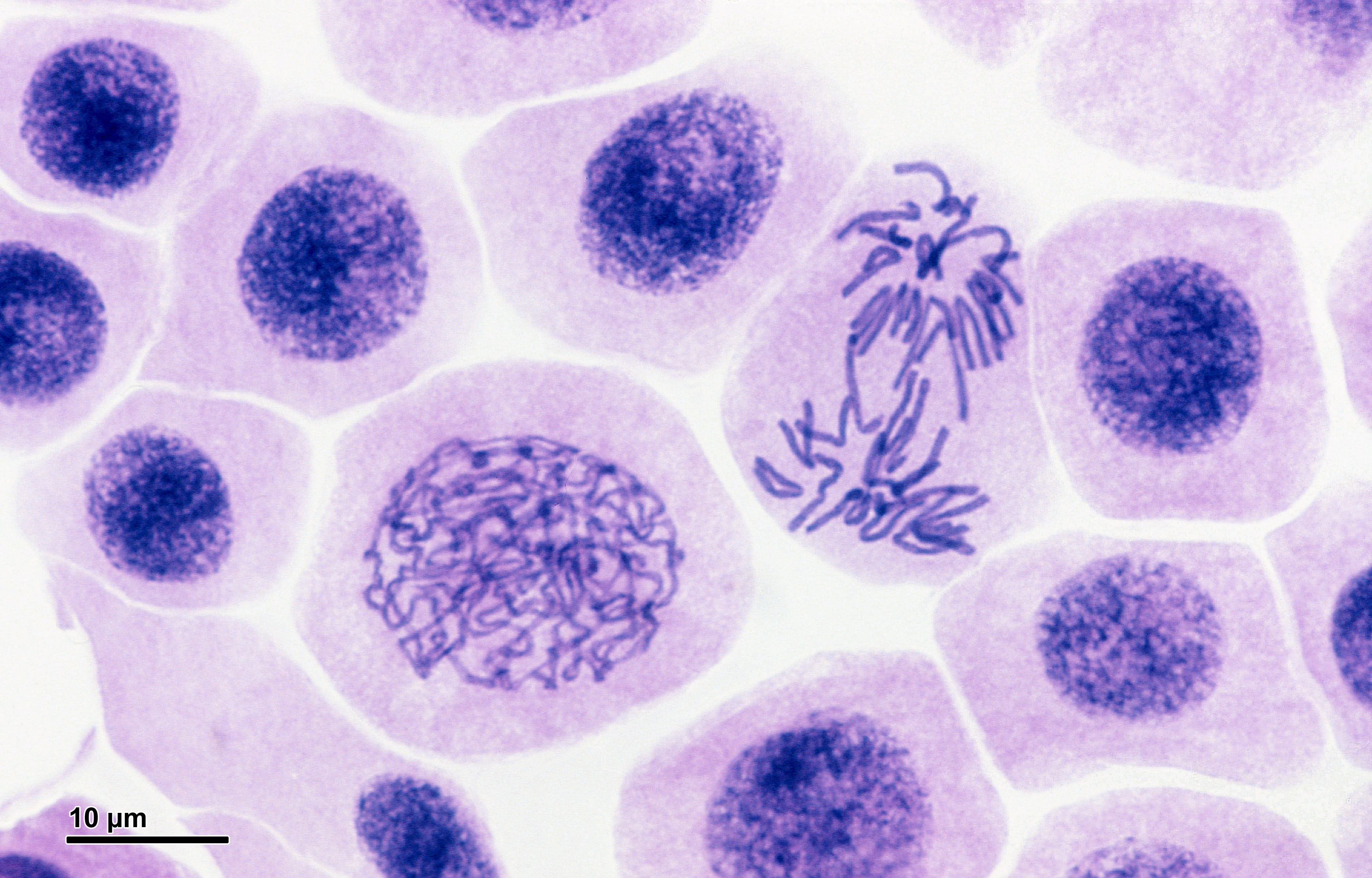
|
| Dr. Josef Reischig, CC BY-SA 3.0 |
| Die dunkel angefärbten Zellkerne der meisten Zellen befinden sich in der Interphase. Etwas links und unterhalb der Bildmitte sieht man einen Zellkern in der Prophase oder Prometaphase. Etwas rechts und oberhalb der Bildmitte sieht man einen Zellkern in der Anaphase. |
| Zwiebelwurzelzellen in Inter-, Pro-, Meta-, Ana- und Telophase |
|---|

|
| Dr. Josef Reischig, CC BY-SA 3.0 |
| Die dunkel angefärbten Zellkerne der meisten Zellen befinden sich in der Interphase. Jeweils eine Zelle befindet sich in der Prophase (links oben), Metaphase (rechts oben), Anaphase (links unten) bzw. Telophase (rechts Mitte). |
Das folgende Bild zeigt Mitose und Cytokinese als kontinuierliche Entwicklung.
| Confocal-lichtmikroskopisches Bild vom letzten Stadium einer Mitose. |
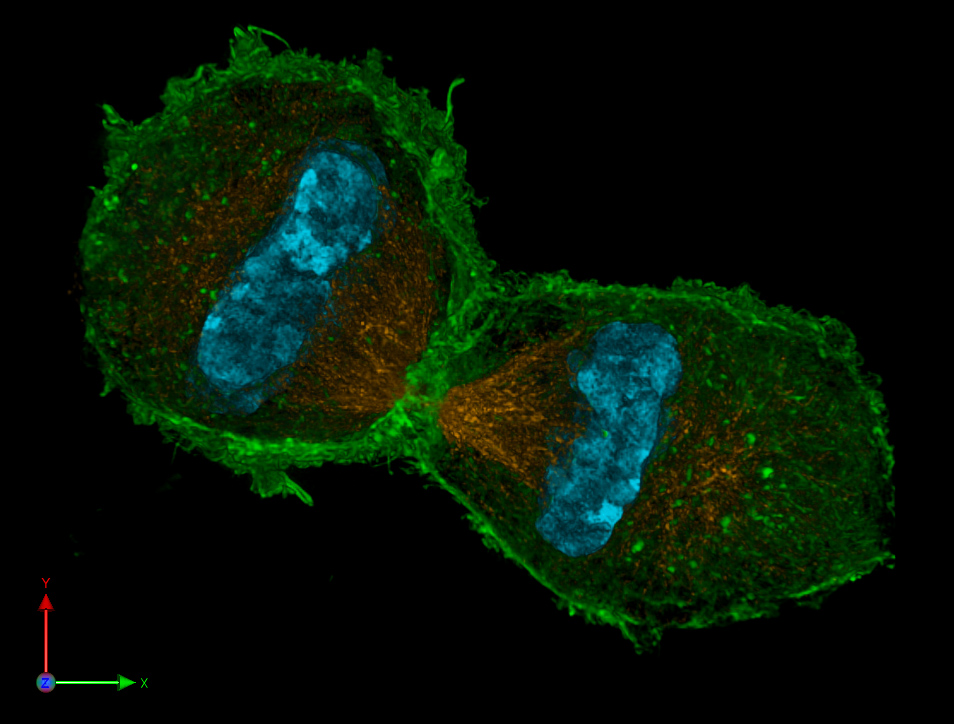
|
| Lothar Schermelleh, CC BY-SA 3.0 |
| Das Bild zeigt in den gerade entstehenden Tochterzellen blau das Chromatin, orange den Spindelapparat und grün das Aktin im Zytoskelett |
Die folgende vereinfachte Animation soll Mitose und Cytokinese als kontinuierliche Entwicklung zeigen.
Die Animation funktioniert unter MS-Windows mit den Browsern Firefox, Opera und Google Chrome, während der Internet Explorer von Microsoft die SMIL-Anweisungen immer noch nicht versteht. Auf dem iPad muss die Seite vor jeder Wiederholung neu aufgebaut werden.
Transkription und DNA-Replikation funktionieren am besten, wenn die DNA möglichst entspannt ist. Schließlich müssen die DNA-Doppelstränge getrennt und um neue Gegenstränge ergänzt werden. Man nennt die Phase der DNA-Replikation auch Synthese-Phase und in ihr kann man die Chromosomen lichtmikroskopisch nicht sehen. Nach der DNA-Replikation enthält jedes Chromosom zwei DNA-Doppelstränge. Jeder der beiden DNA-Doppelstränge ist um spezielle Proteine gewickelt und man nennt diese aus Proteinen und DNA bestehende Struktur ein Chromatid. An einer Centromer genannten Stelle sind die beiden Chromatiden eines 2-Chromatiden-Chromosoms miteinander verbunden. Damit jeder der beiden neuen Zellkerne von jedem Chromosom eine Kopie erhält, müssen in jedem Chromosom die beiden Chromatiden voneinander getrennt und in unterschiedliche Hälften der Zelle gezogen werden. Man kann aber sehr lange DNA-Fäden sehr schlecht transportieren. Vor dem Transport werden deshalb die Chromatiden extrem dicht verknäuelt, sodass sie dick, kurz und dadurch auch sichtbar werden. Danach werden zunächst alle Chromosomen in eine Ebene in der Zellmitte transportiert. Dann werden die beiden Chromatiden jedes Chromosoms getrennt und in entgegengesetzte Richtungen gezogen. Schließlich bildet das endoplasmatische Retikulum in jeder Zellhälfte eine neue Kernhülle um die jetzt nur noch aus jeweils einem Chromatid bestehenden Chromosomen. Diese entknäueln sich jetzt wieder und werden dadurch wieder quasi unsichtbar.
Meiose oder Reduktionsteilung nennt man eine besondere Form der Zellteilung, bei der in zwei Teilungsschritten aus einer diploiden Zelle vier haploide Geschlechtszellen entstehen. Man kann die Meiose grob in die beiden Abschnitte Meiose I und Meiose II unterteilen.
Die meisten Quellen gehen überhaupt nicht auf diese Frage ein, aber der BioCoach des Pearson Verlages erklärt, dass die 2-Chromatiden-Chromosomen in vielen, aber nicht in allen Zellen nach der ersten meiotischen Kernteilung dekondensieren.
Um die Meiose verstehen zu können, sollte man 2 Dinge wissen:
Die Verteilung der ursprünglich mütterlichen und väterlichen Chromosomen erfolgt zufällig und zusätzlich kommt es noch zum Austausch von Chromosomenteilen durch das sogenannte Crossing-over, sodaß jede Geschlechtszelle eine einzigartige Mischung der elterlichen Gene enthält.
|
Schema der Stadien der Meiose mit Crossing over
|
|---|
|
Die Meiose kommt nur bei Lebewesen vor, deren noch lebende Zellen einen Zellkern haben. Diese Lebewesen heißen Eukaryoten. Eukaryoten setzten die Meiose nur für die Herstellung von Geschlechtszellen ein. Der Zweck der Meiose ist die Halbierung des in normalen Körperzellen doppelten (diploiden) Chromosomensatzes. Denn Geschlechtszellen dürfen von jedem Chromosom nur ein Exemplar enthalten, damit sich die Zahl der Chromosomen in den Körperzellen nicht bei jeder sexuellen Fortpflanzung verdoppelt. Die Meiose beginnt mit Chromosomenpaaren aus jeweils zwei Chromosomen, die normalerweise Varianten der gleichen Gene in der selben Reihenfolge enthalten. Sich so entsprechende Chromosomen nennt man homologe Chromosomen. Dabei stammt jeweils ein Chromosom von der Mutter und das andere vom Vater. Und weil schon vor der Meiose jedes Chromosom kopiert oder genauer verdoppelt wurde, besteht jedes Chromosom aus zwei Kopien, die man Chromatiden nennt. Deshalb nennt man die Chromosomen vor der Meiose 2-Chromatiden-Chromosomen. |

|
| anonym, CC BY-SA 4.0 |
|
Das Schema oben beginnt mit drei Paaren homologer Chromosomen mit je zwei Chromatiden. Dabei stammen die blauen Chromosomen vom einen und die roten vom anderen Elternteil. Gelb gezeichnet sind die für den Transport der Chromosomen benötigten Teile des Zytoskeletts. Das Schema unterteilt die Meiose in zwei Zellteilungen (1. Reduktionsteilung, 2. Äquationsteilung genannte normale Mitosen) mit insgesamt acht Phasen:
|
Die Meiose II entspricht der normalen Mitose mit 1-Chromatid-Chromosomen.
Die Animation funktioniert unter MS-Windows mit den Browsern Firefox, Opera und Google Chrome, während der Internet Explorer von Microsoft die SMIL-Anweisungen immer noch nicht versteht. Auf dem iPad muss die Seite vor jeder Wiederholung neu aufgebaut werden.

